

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Breast cancer is the most common malignant tumor in women worldwide, and its incidence has increased tremendously in the past several decades [1]. Despite earlier diagnosis and advances in treatment, breast cancer is a heterogeneous disease with a nearly 30% mortality rate because of disease progression. Cancer invasion and metastasis are pathol-ogical hallmarks of breast cancer and indicate poor prognosis [2].
Matrix metalloproteinases (MMPs) are key enzymes for invasion and metastasis, which are the major causes of mortality in breast cancer patients [3]. Among them, MMP-9 expression is linked with increased metastasis in various tumors, including the brain, prostate, and breast [4]. 12-O-tetradecanoylphorbol-13-acetate (TPA) induces MMP expression by activating transcription factors such as nuclear factor κB (NF-κB) and activator protein-1 (AP-1) [5]. Induction of MMP-9 is critical for TPA-induced breast tumor cell migration and invasion [6]. Thus, controlling MMP-9 in tumors is considered a promising strategy for inhibiting metastasis and improving the survival of breast cancer patients.
Peroxisome proliferator-activated receptors (PPARs) are nuclear receptors that function as transcription factors to regulate the expression of genes involved in lipid oxidation, transport, and storage [7]. Among the three PPAR isoforms (α, β, and γ), PPARγ forms heterodimer with the retinoid X receptor and optimal activation of the heterodimer requires both ligands [8]. The natural ligands of PPARγ are hydroxyoctadecanoic acids and the cyclopentane prostaglandin 15-deoxy-Δ-12, 14 prostaglandin J2; synthetic ligands include troglitazone, rosiglitazone, and ciglitazone [9].
Because abnormal energy metabolism and biological disorder are characteristics of tumors, metabolic control may be effective for cancer management [10]. Normal and cancer cells show large differences in energy metabolism [11]. In addition, increased aerobic glycolysis and elevated oxidative stress are important biochemical features of cancer cells according to the Warburg hypothesis [12]. Although sex and age are key risk factors for breast cancer, a high-fat diet and obesity are also associated with its development [13]. Obesity and associated adipocyte pathology were recently suggested to occur because of downregulation of PPARγ activity [9]. These data as well as the association of obesity with increased breast cancer risk suggest a role for PPARγ activators in the prevention or treatment of breast cancer. We tested a PPARγ activator as a new therapeutic agent for breast cancer. In this study, the effects of troglitazone on MMP-9 expression and invasion in MCF-7 cells were evaluated.
METHODS
Cell culture and reagents
Human breast cancer MCF-7 cell lines were obtained from the American Type Culture Collection (Manassas, USA). Cells were grown in high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics (10,000 U/mL penicillin and 10,000 µg/mL streptomycin) at 37℃ in a 5% CO2 incubator. Troglitazone was purchased from ALEXIS Biochemicals (Lausen, Switzerland) and TPA, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-tetrazolium bromide assay (MTT), and β-actin antibody were obtained from Sigma-Aldrich (St. Louis, USA). Phosphorylated (p)-c-Jun, nuclear factor of κ light polypeptide gene enhancer in B cells inhibitor (p-IκB) α, IκB kinase (p-IKK) α/β, c-Jun N-terminal kinases (JNK), p-JNK, extracellular signal-regulated kinases (ERK), p-ERK, and p-protein kinase B (AKT) antibodies were purchased from Cell Signaling Technology (Danvers, USA). MMP-9, p50, p65, proliferating cell nuclear antigen, IκBα, AKT, and horseradish peroxidase (HRP)-conjugated IgG antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, USA). The mitogen-activated protein kinase (MAPK) inhibitors PD98059 (ERK inhibitor) and SP600125 (JNK inhibitor) and phosphoinositol-3-kinase (PI3K) inhibitors LY294002 and wortmannin were purchased from Merck Millipore (Billerica, USA). NF-κB and AP-1 consensus oligonucleotides were obtained from Promega (Madison, USA). [α-32P] deoxycytidine triphosphate (dCTP) was purchased from Amersham (Buckinghamshire, UK) and DMEM, FBS, and phosphate-buffered saline (PBS) were obtained from Gibco-BRL (Grand Island, USA).
MTT assay
Cell viability was monitored in an MTT assay. MCF-7 cells were seeded in a 96-well plate and incubated at 37℃ for 24 hours to allow attachment. The attached cells were either untreated or treated with different concentrations of troglitazone at 37℃. After inoculation for 24 hours, the cells were washed with PBS before the addition of MTT (0.5 mg/mL) and incubated for 30 minutes at 37℃. Dimethyl sulfoxide was added to dissolve the formazan crystals and absorbance at 570 nm was measured with a microplate reader (Bio-Rad, Hercules, USA).
Nuclear extraction
MCF-7 cells were treated with troglitazone in the presence or absence of TPA for 4 hours. Subsequently, the cells were washed and scraped into PBS (pH 7.5), and pelleted for 3 minutes. Cytoplasmic and nuclear extracts were obtained using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Pierce Biotechnology, Rockford, USA).
Western blotting
After treatment with troglitazone, cells were incubated with TPA for 24 hours at 37℃. These cells were lysed with M-PER Mammalian Protein Extraction Reagent (Pierce Biotechnology). The cell lysates were loaded and resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and proteins were transferred to Hybond™-polyvinylidene fluoride membranes (GE Healthcare Life Sciences, Little Chalfont, UK). Membrane blocking was performed using 2% bovine serum albumin or 5% skim milk and followed by the incubation with primary antibody overnight at 4℃. HRP-conjugated IgG (1:2,000 dilution) was used as the secondary antibody for 1 hour at 4℃. A Fujifilm image analyzer (Tokyo, Japan) was used to determine protein levels. Immunoreactive signals were visualized using the western chemiluminescent HRP substrate (Sigma-Aldrich).
Zymography assay
Conditioned media were collected after 24 hours of treatment, mixed with non-reducing sample buffer, and separated by PAGE containing 0.1% gelatin. Gels were washed for 30 minutes at 18℃ to 23℃ (room temperature) with 2.5% Triton X-100 and then incubated for 16 hours in developing buffer at 37℃. Gels were stained with 0.25% Coomassie brilliant blue (40% methanol and 7% acetic acid) for 30 minutes and photographed using a Fujifilm image analyzer. For densitometric analysis, we used Multi Gauge Image Analysis software (Fujifilm).
Real-time polymerase chain reaction
Total RNA was isolated from cells with TRIzol reagent (Takara, Shiga, Japan) according to the manufacturer's protocol and extracted using FastPure™ RNA Kits (Takara). cDNA was synthesized using a PrimeScript™ RT Reagent Kits (Takara) at 37℃ for 15 minutes and 85℃ for 5 seconds. MMP-9 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA were determined by real-time polymerase chain reaction (PCR) using the ABI PRISM 7900 sequence detection system and SYBR® Green PDR Master Mix (Applied Biosystems, Foster City, USA). Primer sequences for MMP-9 (NM 004994) and GAPDH (NM002046) were as follows: MMP-9: forward, CCTGGAGACCTGAGAACCAATCT; reverse, CCACCCGAGTGTAACCATAGC; GAPDH: forward, ATGGAAATCCCATCACCATCTT; reverse, CGCCCCACTTGATTTTGG. The results were normalized to the expression of GAPDH for variation control in mRNA concentration. The relative expression of each gene was calculated and normalized using the 2−ΔΔCt method. These experiments were repeated at least three times independently.
Luciferase assay
MCF-7 cells were inoculated into 24-well plates. Cells were transfected with NF-κB or AP-1 reporter plasmids using Lipofectamine 2000 (Invitrogen, Carlsbad, USA) at 70%–80% confluence. Transfected cells were pretreated with troglitazone at the indicated concentrations for 1 hour and then with 100 nM TPA. Dual Luciferase Reporter Assay Kits (Promega) and Lumat LB 9507 luminometer (EG&G Berthold, Gaithersburg, USA) were used to detect luciferase activity.
Invasion assay
For the invasion assay, we used 8-µm pore sized 24-well chambers coated with Matrigel (BD Biosciences, Franklin Lakes, USA). Cells were added to the upper chamber and lower compartment containing conditioned medium. After incubation for 24 hours, cells in the upper chamber were removed using cotton swabs. Migrated cells were fixed and stained with Toluidine blue. Invading cells were counted in five random areas of the membrane with a light microscope.
Electrophoretic mobility shift assay
Nuclear extracts were prepared as described previously. Oligonucleotides containing the NF-κB (5′CCGGTTAACAGAGGGGGCTTTCCGAG-3′) or AP-1 (5′CGCTTGATG AGTCAGCCGGAA-3′) binding sites were used as probes (Promega). Complementary strands were labeled with [α-32P]dCTP. Labeled oligonucleotides, 10 µg nuclear extracts and binding buffer were incubated for 30 minutes at room temperature. Reaction products were analyzed using 4% PAGE in 0.5×Tris-borate buffer. A 50-fold excess of NF-κB or AP-1 oligonucleotide was used a control to confirm the specific bands.
Statistical analysis
Data from three or more independent experiments were presented as the mean±standard error of the mean. Statistical analyses were performed by analysis of variance and Duncan's tests using the Microsoft 2010 Excel program (Redmond, USA). In the experiment, p<0.05 was considered as statistically significant.
RESULTS
Effect of troglitazone on MMP-9 expression in MCF-7 cells
MCF-7 cells were treated with troglitazone (0–50 µM) for 24 hours and toxicity was analyzed using an established MTT assay. Troglitazone treatment did not significantly change MCF-7 cell viability (Figure 1A). Therefore, we used in a nontoxic concentration (25 and 50 µM) of troglitazone. We also found that troglitazone induced PPARγ expression in MCF-7 cells (Figure 1B).
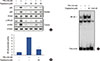
Zymography showed that TPA increased MMP-9 expression in MCF-7 cells and that troglitazone dose-dependently suppressed TPA-induced MMP-9 expression. However, MMP-2 expression was not affected by troglitazone. Western blotting and real-time PCR revealed that troglitazone suppressed TPA-induced MMP-9 expression at both protein and mRNA levels (Figure 2A and 2B). The luciferase assay showed that treatment of MCF-7 cells with troglitazone suppressed TPA-induced MMP-9 promoter activity (Figure 2C). To address whether troglitazone had inhibitory effects via a PPARγ-dependent or PPARγ-independent pathway, we confirmed that suppression of MMP-9 expression after pretreatment with troglitazone was recovered by treatment of MCF-7 cells with the PPARγ antagonist GW9662 (Figure 2D). In addition GW9662 treatment did not affect TPA-induced MMP-9 expression.
Effects of troglitazone on ERK/JNK and PI3K/AKT activation in MCF-7 cells
To investigate the effect of troglitazone on ERK/JNK and PI3K/AKT activity, we performed zymography and western blotting. MCF-7 cells were pretreated with inhibitors of ERK (PD98059), JNK (SP600125), or PI3K (LY294002 and wortmannin). The inhibition of ERK, JNK, or PI3K blocked TPA-induced MMP-9 protein expression in MCF-7 cells (Figure 3A and 3B). In addition, TPA significantly enhanced the phosphorylation of ERK, JNK and AKT in MCF-7 cells; this effect was blocked by troglitazone (Figure 3C). MCF-7 cells were also pretreated with an inhibitor of NF-κB (Bay 11-7092) and AP-1 (curcumin). The inhibition of NF-κB and AP-1 suppressed TPA-induced MMP-9 expression in MCF-7 cells (Figure 3D).
Effect of troglitazone on TPA-induced NF-κB and AP-1 activation in MCF-7 cells
To understand the mechanism by which troglitazone inhibited MMP-9 transcription, we tested whether troglitazone inhibited NF-κB and AP-1 activation in MCF-7 cells after TPA stimulation. Troglitazone inhibited nuclear translocation of NF-κB p65/p50 and phosphorylation of IKKα/β and IκBα as determined by western blotting of MCF-7 cells (Figure 4A). The luciferase assay revealed that troglitazone inhibited the promoter activity of TPA-induced NF-κB (Figure 4B). In addition, electrophoretic mobility shift assay showed that troglitazone significantly inhibited TPA-induced NF-κB DNA binding (Figure 4C). Troglitazone inhibited phosphorylation of c-Jun and c-Fos and the promoter activities of TPA-induced AP-1 (Figure 5A and 5B). Troglitazone also inhibited TPA-induced AP-1 DNA binding (Figure 5C).
Effect of troglitazone on in vitro invasion of TPA-induced MCF-7 cells
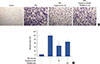
A Matrigel invasion assay was used to elucidate the inhibition by troglitazone of invasion on MCF-7 cells. A troglitazone concentration of 50 µM effectively inhibited the invasion of MCF-7 cells by approximately 55% compared to untreated control cells. However, inhibition of only 30% was observed with troglitazone and GW9662, indicating that troglitazone suppresses the invasive potential of MCF-7 cells via PPARγ activation (Figure 6).
DISCUSSION
We found that troglitazone, a synthetic PPARγ ligand, blocks TPA-induced MMP-9 expression and invasion of MCF-7 cells through a PPARγ-dependent mechanism. Troglitazone blocked cancer cell invasion by suppressing NF-κB/AP-1-mediated MMP-9 expression. Thus, troglitazone may be developed as a novel therapeutics to prevent breast cancer invasion.
PPARγ ligands are potential targets for the prevention and treatment of various human cancers [14]. PPARγ ligands inhibit proliferation and induce apoptosis of breast cancer cells [1516], indicating their potential for breast cancer treatment and prevention. PPARγ protein is expressed in breast cancer cell lines including MCF-7 and MDA-MB-231 cells [17]. Troglitazone induces PPARγ expression in many tumors including breast, brain, and liver [18]. We found that troglitazone induces PPARγ-dependent G1 arrest and apoptosis of breast cancer cells [19], indicating that PPARγ-regulates genes that are pivotal in breast carcinogenesis.
NF-κB and AP-1 are well-known, major transcriptional factors that regulate MMP-9 expression in cancer cells [20]. MAPK and PI3K/AKT are signaling transducers upstream of NF-κB and AP-1; their activation stimulates the growth and invasion of cancer cells [212223]. Our data suggest that troglitazone inhibits the NF-κB, AP-1, MAPK, and PI3K/AKT signaling pathways in MCF-7 cells. This finding is important for understanding breast cancer progression, as these signaling pathways increase tumor cell invasiveness because of increased MMP-9 expression [24]. We found that troglitazone inhibited MMP-9 expression in MCF-7 cells and suppressed the invasion of breast cancer cells. Troglitazone inhibited tumor invasion by suppressing NF-κB-, AP-1-, MAPK-, and PI3K/AKT-dependent MMP-9 expression and invasion of MCF-7 cells. In addition, suppression of MMP-9 expression by troglitazone in MCF-7 cells was recovered by GW9662, an inhibitor of PPARγ, indicating that the troglitazone inhibitory effects involve a PPARγ-dependent pathway. Thus, we suggest that troglitazone involves PPARγ-dependent inhibitory mechanisms affecting MMP-9 expression in various cancer cell lines.
In conclusion, we investigated whether MMP-9 expression and invasion of breast cancer cells is modulated by PPARγ ligands. MCF-7 cells treated with troglitazone displayed inhibited MMP-9 expression, which plays a critical role in cancer invasion. Moreover, troglitazone caused a PPARγ-dependent decrease of MMP-9 expression in MCF-7 cells. Our results suggest that troglitazone affects estrogen receptor-positive breast cancer metastasis and shows potential for the prevention and treatment of estrogen receptor-positive breast cancer. However, the therapeutic effects of troglitazone on estrogen receptor-negative breast cancer requires further analysis.
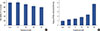



 XML Download
XML Download