

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Berberine (BBR) is an isoquinoline alkaloid isolated from the Chinese herb Coptis chinensis (Huanglian) and is a promising component of medicinal plants [1]. BBR represses tumor progression by suppressing abnormal cell proliferation, arresting the cell cycle, and inducing apoptosis in breast cancer and osteosarcoma cells [23]. Furthermore, BBR suppresses the metastatic potential of breast cancer cells through downregulation of matrix metalloproteinase (MMP)-1 and MMP-9 [45]. BBR regulates various signaling pathways including mitogen-activated protein kinases (MAPKs), c-Jun N-terminal protein kinase (JNK), and nuclear factor κB (NF-κB) in a variety of cancer cells [678]. We also reported that BBR suppresses 12-O-tetradecanoylphorbol-13-acetate-induced vascular endothelial growth factor expression through inhibition of the phosphatidylinositol-4,5-bisphosphate 3-kinase/protein kinase B (AKT) pathway in breast cancer cells [9]. Here, we explored the regulatory mechanism of BBR on fibronectin (FN) expression in triple-negative breast cancer (TNBC) cells.
FN is one of the most abundant extracellular matrix glycoproteins and is implicated in physiological and pathological processes such as cell differentiation, invasion, migration, and oncogenic transformation [1011]. FN mRNA and protein expression is dramatically increased in the stroma of breast tumors compared with normal adult breast tissues [12]. The cell adhesive region of FN comprises at least two minimal amino acid sequences: an Arg-Gly-Asp (RGD) sequence and a Pro-His-Ser-Arg-Asn sequence [1314]. The RGD site of FN binds to specific cell surface receptors called integrins, and the resulting complex plays a crucial role in the progression of breast cancer metastasis [13]. Administration of synthetic peptides containing the RGD sequence can inhibit FN cell adhesion function and metastatic potential in breast cancer models [15].
This study aimed to investigate the mechanism by which BBR inhibits FN expression in TNBC cells. We examined the clinical relevance of FN expression in breast cancer patients and demonstrated that BBR completely suppresses the basal levels of FN expression in TNBC cells through inhibition of activator protein (AP)-1 activity.
METHODS
Reagents
Dulbecco's modified Eagle's medium (DMEM), Roswell Park Memorial Institute (RPMI) 1640, and antibiotics were purchased from Life Technologies (Rockville, USA). Fetal bovine serum (FBS) was purchased from Hyclone (Logan, USA). BBR was purchased from Sigma (St. Louis, USA). SR11302 was purchased from Tocris (Ellisville, USA). The secondary horseradish peroxidase (HRP)-conjugated antibody and mouse monoclonal anti–β-actin antibody were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, USA). Rabbit monoclonal anti-FN antibodies to phospho (p)-c-Jun, total (t)-c-Jun, and c-Fos were purchased from AbCam (Cambridge, UK). The enhanced chemiluminescence (ECL) prime reagents were from Amersham (Buckinghamshire, UK).
Analysis of public database
Expression data were downloaded from a public database (Kaplan-Meier plotter database; http://kmplot.com/breast) [16]. The clinical relevance of FN mRNA expression in breast cancer patients was analyzed by Kaplan-Meier survival plots. The hazard ratio with 95% confidence interval and log-rank p-values were calculated.
Cell culture and drug treatment
MDA-MB453, BT20, BT549, MDA-MB231, Hs578T, and MDA-MB157 human breast cancer cells were grown in a humidified atmosphere of 95% air and 5% CO2 at 37°C in DMEM supplemented with 10% FBS, 2 mM glutamine, 100 IU/mL penicillin, and 100 µg/mL streptomycin. BT474, T47D, ZR75-1, SKBR3, and HCC1143 human breast cancer cells were grown in RPMI1640 media under the same conditions.
For drug treatment, cells were maintained in culture medium without FBS for 24 hours and then treated with the indicated concentrations of BBR or SR11302 for 24 hours in fresh serum-free media.
Real-time polymerase chain reaction
Total RNA was extracted from cells using the TRIzol reagent (Invitrogen, Carlsbad, USA), according to the manufacturer's instructions. Isolated RNA samples were then used for reverse transcriptase-polymerase chain reaction (RT-PCR). Samples (1 µg of total RNA) were reverse-transcribed into cDNA in 20-µL reaction volumes using a first-strand cDNA synthesis kit for RT-PCR, according to the manufacturer's instructions (MBI Fermentas, Hanover, USA).
Gene expression was quantified by real-time PCR using a SensiMix SYBR Kit (Bioline Ltd., London, UK) with 100 ng of cDNA per reaction. The sequences of primer sets used for this analysis are as follows: human FN (forward, 5′-CCA CCC CCA TAA GGC ATA GG-3′; reverse, 5′-GTA GGG GTC AAA GCA CGA GTC ATC-3′) and glyceraldehyde-3-phosphate dehydrogenase as an internal control (forward, 5′-ATT GTT GCC ATC AAT GAC CC-3′; reverse, 5′-AGT AGA GGC AGG GAT GAT GT-3′). An annealing temperature of 60°C was used for all primers. PCR was performed in a standard 384-well plate format on an ABI 7900HT real-time PCR detection system. For data analysis, the raw threshold cycle (CT) value was first normalized to the housekeeping gene for each sample in order to obtain the ΔCT. The normalized ΔCT was then calibrated to the control cell samples to obtain the ΔΔCT.
Western blotting
Cell culture media (supernatants) and cell lysates were used in immunoblot analysis for FN, c-Jun, c-Fos, and β-actin. The protein samples were boiled for 5 minutes in Laemmli sample buffer and electrophoresed in 8% (FN or β-actin) or 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The separated proteins were transferred to polyvinylidene fluoride membranes, and the membranes were blocked with 10% skim milk in tris-buffered saline (TBS) with 0.01% Tween-20 for 15 minutes. The blots were incubated overnight with antibodies against FN, c-Jun, c-Fos, and β-actin in 1% TBS/T buffer (0.01% Tween-20 in TBS) at 4°C. The blots were then washed 3–4 times in TBS with 0.01% Tween-20 and subsequently incubated with anti-rabbit peroxidase-conjugated antibody (1/2,000 dilution) in TBS/T buffer. After 1 hour of incubation at room temperature, the blots were washed three times, and developed using ECL prime reagents.
Cell adhesion and spreading assay
MDA-MB231 and Hs578T cells were grown to approximately 80% confluence in DMEM containing 10% FBS, detached, and then suspended in serum-free medium. The cell suspensions (5×104 cells in 250 µL) were seeded into each well of 24-well plates precoated with 100 ng/mL recombinant human FN (rhFN). After incubation at 37°C for 3 hours, nonadherent cells were removed by washing with phosphatebuffered saline. Cell spreading was analyzed using a CK40 inverted microscope (Olympus, Tokyo, Japan). Adherent cells were fixed in 4% formaldehyde and stained with 0.2% crystal violet in 10% ethanol. The stained cells were then dissolved in 2% SDS, and absorbance at 570 nm was measured using a spectrophotometer (Spectra Max 190; Molecular Devices, Sunnyvale, USA).
Statistical analysis
Statistical significance was determined using the Student t-test. Data are presented as mean±standard error of the mean. All quoted p-values are two-tailed, and differences were considered significant for p<0.05. Microsoft Excel was used for statistical analyses (Microsoft, Redmond, USA).
RESULTS
Aberrant FN expression is associated with poor prognosis in breast cancer patients
We analyzed whether FN expression is associated with survival in breast cancer patients using the Kaplan-Meier plotter database (http://kmplot.com/breast) [16]. Abnormal FN expression was associated with poor prognosis for relapse-free survival (p=0.006) (Figure 1A) and overall survival (p = 0.003) (Figure 1B) in breast cancer patients.
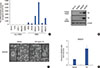
Next, we investigated FN mRNA expression levels in a variety of breast cancer cells. The clinicopathological features of breast cancer cell lines were determined from the American Type Culture Collection website (https://www.atcc.org). We also classified cells as non-TNBC (BT474, T47D, ZR75-1, MDA-MB453, and SKBR3) and TNBC (BT20, BT549, MDA-MB231, Hs578T, HCC1143, and MDA-MB157) to compare the levels of FN mRNA expression. As shown in Figure 2A, FN mRNA expression was significantly elevated in TNBC cells compared with non-TNBC cells. The FN mRNA expression was 2587.8-fold (MDA-MB231 cells) and 6555.5-fold (Hs578T cells) greater than the control levels (BT474 cells) (Figure 2A). We also confirmed the level of FN protein expression in non-TNBC cells (BT474 and SKBR3) and TNBC cells (MDA-MB231 and Hs578T) using culture media and whole cell lysates. As expected, FN protein levels were dramatically increased in TNBC cells (Figure 2B). In a previous study, we reported that upregulation of FN by human epidermal growth factor receptor 2 (HER2) overexpression augments cell adhesion and invasion in breast cancer cells [15]. We therefore analyzed the morphological changes induced by treatment with 100 ng/mL rhFN. Extension of protrusions was increased in rhFN-treated cells compared with the control cells (vehicle-treated) (Figure 2C). In addition, the adhesion rates of MDA-MB231 cells were significantly promoted by rhFN treatment (Figure 2D).
Basal FN expression is suppressed by SR11302 treatment
Next, we investigated whether AP-1 activity is involved in FN expression. Cells in serum-free media were treated with a specific AP-1 inhibitor, SR11302, for 24 hours. As shown in Figure 3A, FN mRNA expression was dramatically decreased by SR11302 in both MDA-MB231 and Hs578T cells by 0.4±0.1-fold and 0.7±0.1-fold, respectively, compared with the control level (Figure 3A). In addition, FN protein expression was dose-dependently decreased by SR11302 in both MDA-MB231 (Figure 3B) and Hs578T cells (Figure 3C). These results demonstrated that FN expression in TNBC cells is regulated through AP-1 activity.
Basal FN expression is suppressed by berberine treatment
We then examined the effect of BBR on FN expression in TNBC cells. After serum starvation for 24 hours, cells were treated with the indicated concentrations of BBR for 24 hours in serum-free media. The chemical structure of BBR is represented in Figure 4A. After 24 hours, we harvested conditioned culture media and prepared whole cell lysates for detection of FN mRNA and protein expression. As shown in Figure 4B, FN mRNA expression was decreased upon treatment with 50 µM BBR. In particular, FN mRNA expression was significantly decreased to 0.25±0.04-fold of the control level in MDA-MB231 cells. Furthermore, FN protein expression was dosedependently decreased by BBR treatment in both MDA-MB231 (Figure 4C) and Hs578T cells (Figure 4D).
To verify the inhibitory mechanism of BBR on FN expression, we examined the phosphorylation levels of various signaling molecules. We found that phosphorylation of c-Jun, a member of the AP-1 family, was significantly decreased by BBR treatment in Hs578T TNBC cells (Figure 4E); however, the level of c-Fos expression was not altered by BBR (Figure 4E). Thus, we demonstrated that BBR suppresses the FN mRNA and protein expression levels through inhibition of AP-1 activity.
DISCUSSION
FN plays a central role in processes associated with tumor cell proliferation and is highly expressed in malignant breast cancer cells, but not in nonmalignant breast epithelial cells [1718]. The blood concentration of FN shows great promise as a marker for cancer and other diseases [19]. The level of plasma FN expression is directly associated with clinical events such as chemotherapy [1920]. The plasma FN level is reportedly higher in patients with metastatic breast cancer than in those with no detectable disease [19]. In addition, abnormal FN induction within tumor associated fibroblasts triggers tumor cell motility, cancer spread, and metastasis formation [21]. In the present study, we also observed that FN expression is directly associated with relapse-free survival and overall survival in breast cancer patients. Therefore, we propose that suppression of FN expression is one of the most important approaches for breast cancer treatment.
FN can trigger a variety of cellular signaling pathways and thus contribute to tumorigenesis [22]. The RGD site of FN binds to β1 integrin and augments the invasiveness of α5β1-integrin–expressing breast cancer cells [23]. FN-stimulated cell migration and invasion are achieved through activation of focal adhesion kinase in non-small cell lung carcinoma [24]. The basal level of FN expression is increased by HER2 overexpression and subsequently triggers cell adhesion and invasion in breast cancer cells [15]. In addition, upregulation of FN gene expression by sex determining region Y-box 2 (SOX2) significantly increased the invasion of ovarian cancer cells [25]. Our results show that the basal levels of FN mRNA and protein expression are significantly increased in TNBC cells. In addition, the rates of cell adhesion and spreading are also increased by FN treatment in MDA-MB231 TNBC cells. We thus demonstrate that FN expression is associated with characteristics of TNBC cells related to aggressiveness, such as cell invasion, adhesion, and migration.
Regions of the FN promoter contain binding sites for inducible transcription factors such as cAMP-responsive element-binding protein, AP-1, and AP-2 [2627]. Angiotensin II (Ang II) activates FN gene transcription in vascular smooth muscle cells via the DNA binding activity of AP-1 [2628]. In contrast, mutation of AP-1 disrupts nuclear binding and suppresses Ang II-induced transcription in the native FN promoter [26]. In the present study, we explored the pharmacological mechanism of BBR for suppressing FN expression in breast cancer cells. Consistent with the above results, we observed that SR11302, a specific AP-1 inhibitor, dose-dependently decreases the levels of FN mRNA and protein expression in TNBC cells, indicating that AP-1 activity plays an important role in FN expression in TNBC cells.
To date, BBR has been shown to have various pharmacological functions as a traditional medicine or dietary supplement against fungal, bacterial, and viral infections and neoplastic disease [1]. BBR induces cytotoxicity through upregulation of the proapoptotic genes Fas, FasL, p53, and Bax in colon cancer, hepatoma, and gastric carcinoma cells [2930]. BBR suppresses several signaling pathways including MAPKs, JNK, and NF-κB in melanoma and breast cancer cells [678]. Here, we observed that BBR inhibits AP-1 activity via suppression of c-Jun phosphorylation in TNBC cells. We further demonstrated that BBR downregulates the FN mRNA and protein expression in TNBC cells through inhibition of AP-1 activity.
In conclusion, we investigated the mechanism by which BBR regulates FN expression in TNBC cells. Our results showed that abnormal FN expression is associated with poor prognosis in breast cancer patients. In addition, FN mRNA and protein expression levels are higher in TNBC cells than in non-TNBC cells. FN plays an important role in TNBC breast cancer cell adhesion and spreading. Interestingly, BBR decreases FN expression in TNBC cells through inhibition of c-Jun phosphorylation. Our results demonstrate that FN could be a novel therapeutic target in breast cancer, and that BBR might be a promising drug for treatment of TNBC through suppression of FN expression.



 XML Download
XML Download