

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Breast cancer is the most commonly diagnosed neoplasm and the leading cause of cancer mortality among women [1]. It is a heterogeneous disease; predicting its clinical behavior and preparing therapeutic plans are challenging despite various histopathological classifications at diagnosis [2]. Clinicopathological factors such as American Joint Committee on Cancer (AJCC) stage, histological grade, estrogen receptor (ER) and progesterone receptor (PR) expression, and human epidermal growth factor receptor 2 (HER2) gene amplification are currently considered in the prognosis and management of breast cancer [3]. Based on emerging molecular and immunohistochemical methods, breast cancer is classified into four major subtypes: luminal A, luminal B, HER2+, and triple-negative. Luminal B is subdivided according to HER2 status (luminal B HER2− and luminal B HER2+) and Ki-67 labeling index [4].
Signaling by the Wnt family of secreted glycolipoproteins is an ancient and evolutionarily conserved pathway linked to cell proliferation, cell polarity, and fate determination [5]. Wnt7a, a member of the Wnt family, is normally expressed in several organs, including the lung, testis, lymph node, and brain [6]. As an oncogenic autocrine glycoprotein, Wnt7a promotes tumor invasion and distant metastasis with cancer-associated fibroblasts [7]. Interestingly, published data have shown that Wnt7a shows different expression patterns in different types of malignancies [6]. Wnt7a exhibits tumorigenic potential and functions as an oncogene in endometrial and ovarian cancer, but ironically seems to suppress tumors in the cervix, lung, leukemia, kidney, and pleura [891011121314]. Additionally, high Wnt7a expression is associated with a good prognosis in mesothelioma [14] but a poor prognosis in ovarian cancer [9]. Therefore, Wnt7a apparently has antithetical biological behaviors in several cancers.
In several breast cancer studies, only one demonstrated a correlation between Wnt7a expression and worse disease-free survival (DFS) in breast cancer, especially in the basal subtype [7]. Although Wnt7a expression was linked to low DFS rate, whether this association is accurate enough to predict the clinical outcome remains disputable when considering the multifactorial nature of breast cancer development.
Here, we investigated the prognostic validity of Wnt7a expression and analyzed any statistical correlations with clinicopathological parameters, DFS, and overall survival (OS) in patients with invasive ductal carcinoma (IDC) of the breast. Moreover, we evaluated how Wnt7a and survival rate are interrelated according to ER, PR, and HER2 status.
Go to : 
METHODS
Patients and specimens
We collected 299 cases of breast cancer at Hanyang University Hospital between January 2000 and December 2009. Clinicopathological information was obtained from the patients' medical records and pathological reports. Among the patients, 41 were excluded due to unavailability of paraffin blocks or inadequate clinical history. The remaining 258 patients diagnosed with IDC were selected as subjects. The median age was 49 years (range, 27–79 years). Among them, 181 patients underwent modified radical mastectomy, 65 underwent breast-conserving operation, 11 underwent skin-sparing mastectomy, and one underwent a simple mastectomy.
The distribution of tumor (T) and node (N) stages was as follows: T1, 120 (46.5%); T2, 122 (47.3%); T3, 8 (3.1%); T4, 8 (3.1%); and N0, 140 (54.3%); N1, 54 (20.9%); N2, 33 (12.8%); N3, 31 (12.0%). Overall, 90 patients received a combination of chemotherapy and tamoxifen, 53 received tamoxifen only, and 103 received chemotherapy only. Ten patients showed recurrence and 49 patients died during the follow-up. The 1-, 3-, and 5-year mortality rates were 11.6%, 8.2%, and 12.1%, respectively. The recurrence rate at the 1-, 3-, and 5-year follow-ups were 0.8%, 2.5%, and 3%, respectively.
This study, along with the waiver of informed consent, was approved by the Institutional Review Board at Hanyang University Hospital (HYUH 2016-05-009).
Tissue microarray construction and immunohistochemistry
We punched a 2-mm core in the most cellular area of each paraffin-embedded tumor tissue block, and manually transferred the specimen to a recipient block. Tissue microarray (TMA) sections (4-µm thick) were stained using anti-Wnt7a antibody (1:50 dilution; ab183653; Abcam, Cambridge, UK) after deparaffinization, heat-induced antigen retrieval with Bond epitope retrieval solution (Leica BioSystems, Newcastle, UK), and endogenous peroxidase blocking. We used a Bond-Max automated immunostainer (Leica BioSystems). We incubated the sections in the primary antibody for 30 minutes at room temperature and developed the sections using the Bond Polymer Refine Detection kit (Leica BioSystems) and 3,3´-diaminobenzidine tetrahydrochloride as a chromogen. Immunohistochemical staining for ER, PR, and HER2 was performed using the following antibodies (Novocastra Laboratories, Newcastle, UK): monoclonal mouse anti-ER (1:50), monoclonal mouse anti-PR (1:100), and monoclonal mouse anti-c-erbB-2 (1:800).
Interpretation of immunohistochemical staining
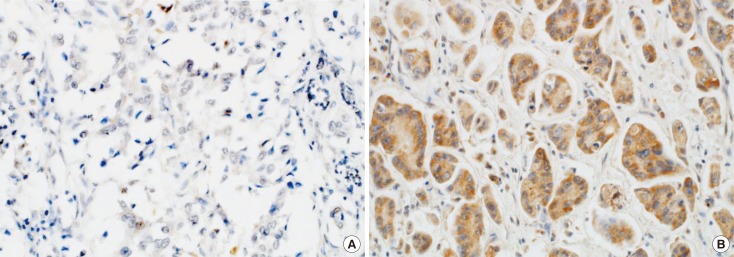
For this study, positive Wnt7a immunostaining was defined as cytoplasmic staining regardless of nuclear staining and graded according to both the intensity and percentage of positively stained tumor cells [815]. Wnt7a staining intensity was scored on a scale of 0 to 3 (0, negative; 1, weak; 2, moderate; 3, strong). The percentage of Wnt7a-positive cells was also classified into four categories: 1 (0%–25%), 2 (26%–50%), 3 (51%–75%), and 4 (76%–100%). The level of Wnt7a staining was evaluated as an immunoreactive score (IRS), which was calculated by multiplying the staining intensity scores and the percentages of positive cells [16]. Based on the receiver operating characteristic (ROC) curve, there was moderate discriminatory power for correlating recurrence rate with Wnt7a expression (area under the ROC, 0.636; sensitivity, 82.7%; specificity, 50%). Wnt7a expression was determined as either negative (IRS <1) or positive (IRS ≥1) (Figure 1). The distribution of the molecular subtypes using immunohistochemical markers was as follows: luminal A (ER+ and/or PR+; HER2−; Ki-67 <14%), 100 (38.8%); luminal B HER2− (ER+ and/or PR+; HER2−; Ki-67 ≥14%), 45 (17.4%); luminal B HER2+ (ER+ and/or PR+; HER2+), 21 (8.1%); HER2+ (ER− and PR−; HER2+), 39 (15.1%); and triple-negative (ER− and PR−; HER2−), 53 (20.5%) [4].
Statistical analysis
Categorical variables were compared using the chi-square test. For ordinal variables, the Mantel-Haenszel method (also called linear-by-linear association) was used to determine trends. Continuous variables were compared using Student t-test. DFS and OS curves were generated using the Kaplan-Meier method and were compared by the log-rank test. Meanwhile, multivariate analysis was performed to identify independent prognostic markers for DFS and OS using a Cox multistep regression model. A p<0.05 was considered statistically significant. All statistical computations were performed with R version 3.2.2 (http://www.R-project.org/).
Go to :
RESULTS
Relationship between Wnt7a expression and clinicopathological parameters and molecular subtypes
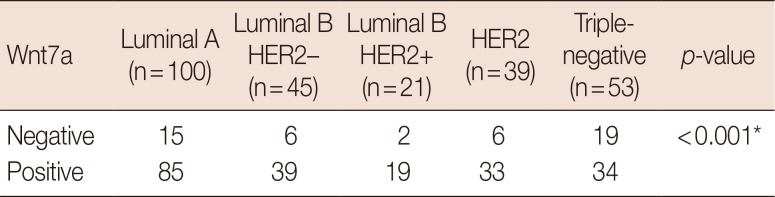
Wnt7a expression was associated with ER positivity (odds ratio [OR], 3.95; 95% confidence interval [CI], 1.99–7.80; p<0.001), which was frequently observed in patients older than 55 years (OR, 3.6; 95% CI, 1.54–8.42; p=0.002). No significant difference existed with other parameters such as tumor size, lymphovascular invasion, perineural invasion, AJCC stage, histological grade, and HER2 expression (Table 1). Regarding molecular subtypes [4], Wnt7a expression was more frequently observed in luminal A, luminal B, and HER2 groups than in the triple-negative group (OR, 3.4; 95% CI, 1.70–6.73; p<0.001) (Table 2, Figure 2).
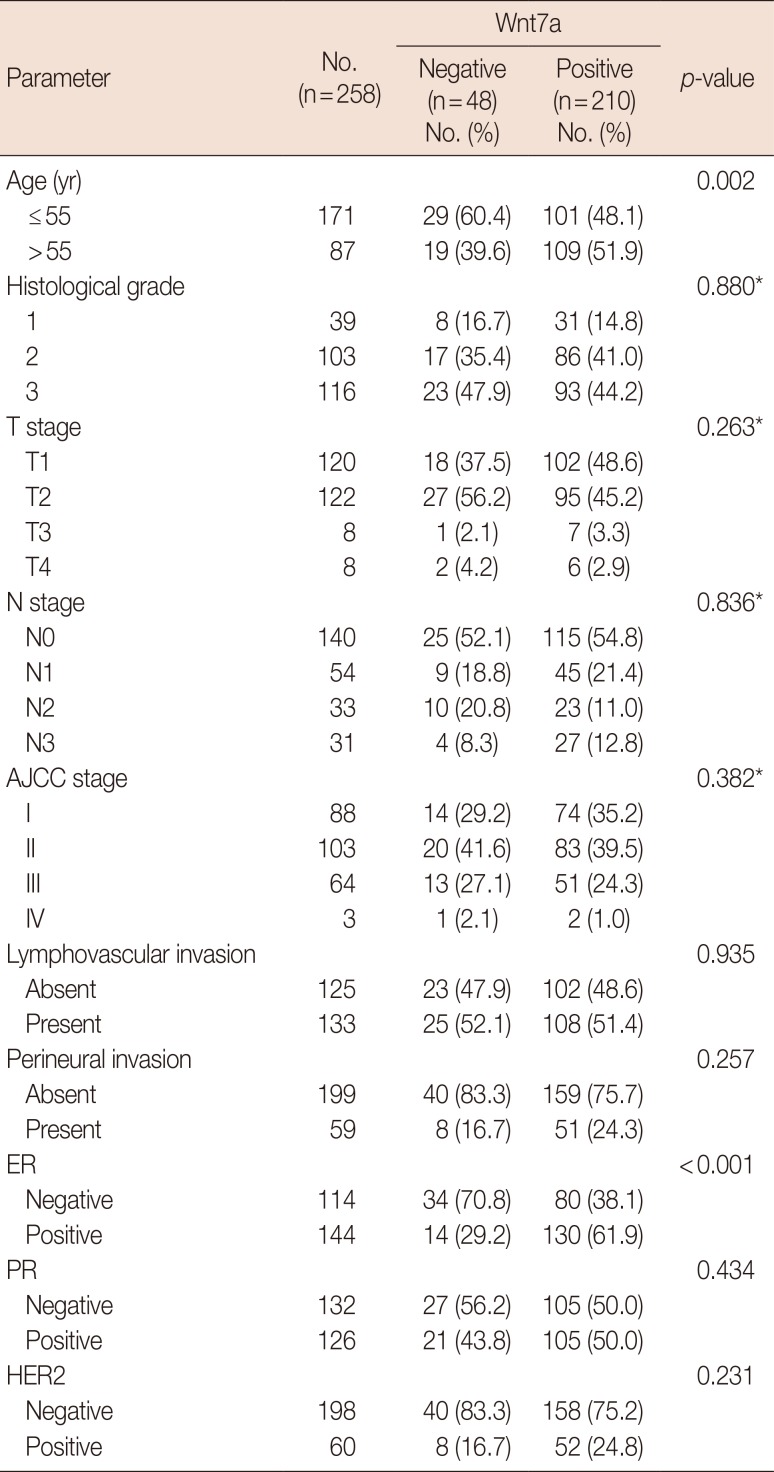
 | Figure 2Expression of Wnt7a according to molecular subtype. Wnt7a negativity is frequently observed in the triple-negative group compared with other groups including luminal A, luminal B human epidermal growth factor receptor 2 (HER2)−, luminal B HER2+, and HER2 (p<0.001). Values in parentheses represent percentage.
|
Table 1
Comparison of clinicopathologic parameters between the Wnt7a-expressing and non-expressing group
![]()
Comparison between patient survival and Wnt7a expression
On univariate survival analysis, Wnt7a negativity was significantly associated with poor DFS (hazard ratio [HR], 4.63; 95% CI, 1.34–15.98; p=0.007), but not with poor OS. Other clinicopathological parameters such as age (HR, 2.46; 95% CI, 1.33–4.54; p=0.004), T stage (HR, 5.41; 95% CI, 1.14–25.56; p=0.033), lymphovascular invasion (HR, 2.16; 95% CI, 1.12–4.19; p=0.021), ER (HR, 1.93; 95% CI, 1.03–3.59; p=0.039), PR (HR, 2.86; 95% CI, 1.43–5.70; p=0.003), and hormone therapy (HR, 4.27; 95% CI, 2.09–8.72; p<0.001) were correlated with OS, whereas only T stage (HR, 5.16; 95% CI, 2.38–11.19; p<0.001) was related to DFS. After adjusting for confounding factors such as T criteria, histological grade, lymphovascular/perineural invasion, hormone therapy, and chemotherapy, Wnt7a negativity (HR, 9.12; 95% CI, 1.80–46.09; p=0.008) and old age (HR, 7.17; 95% CI, 1.19–43.09; p=0.031) were still correlated with worse DFS, whereas advanced T stage (HR, 6.72; 95% CI, 2.89–15.66; p<0.001) and old age (HR, 3.23; 95% CI, 1.62–6.44; p=0.001) were associated with poor OS (Table 3, Figure 3).
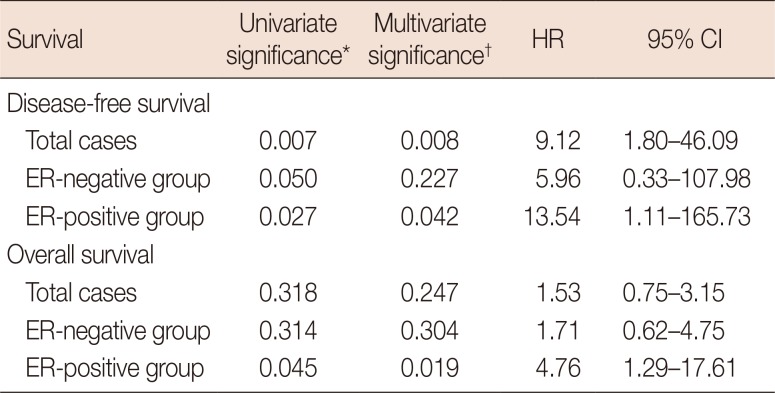
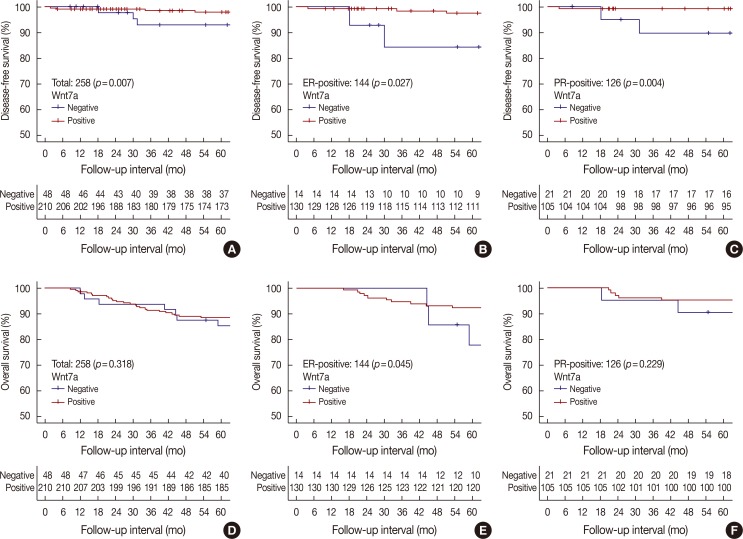 | Figure 3Patients' survival according to Wnt7a expression. Kaplan-Meier curves representing disease-free survival in all cases (A), the estrogen receptor (ER)-positive group (B) and the progesterone receptor (PR)-positive group (C) and overall survival in all cases (D), the ER-positive group (E) and the PR-positive group (F).
|
Table 3
Correlations between disease-free and overall survival and Wnt7a expression
Total cases, 258 patients; ER-negative group, 114 patients; ER-positive group, 144 patients.
HR=hazard ratio; CI=confidence interval; ER=estrogen receptor.
*Log-rank test; †Cox proportional hazard model: adjusted for age (≤55 years vs. >55 years), T stage (1 or 2 vs. 3 or 4), histological grade (1 vs. 2 or 3), lymphovascular/perineural invasion (absence vs. presence), hormone therapy and chemotherapy.
![]()
We also evaluated DFS and OS in the groups based on ER, PR, and HER status. In the ER-positive group, Wnt7a negativity had a significant correlation with poor DFS (multivariate HR, 13.54; 95% CI, 1.11–165.73; p=0.042) and OS (multivariate HR, 4.76; 95% CI, 1.29–17.61; p=0.019). In the PR-positive group, Wnt7a negativity tended to be associated with worse DFS (multivariate HR, 12.29; 95% CI, 0.68–223.49; p=0.090) (Figure 3). Regarding molecular subtypes, Wnt7a negativity was significantly associated with poor DFS in the luminal A group (HR, 6.04; 95% CI, 1.22–29.99; p=0.028). Meanwhile, a significant relationship existed between Wnt7a expression and worse OS in the luminal B HER2+ group (HR, 30.15; 95% CI, 2.67–340.66; p=0.006). However, no statistical significance was shown on multivariate analyses.
Go to :
DISCUSSION
We showed here that Wnt7a expression was correlated with several clinicopathological factors including old age, ER positivity, and DFS in 258 patients with IDC. Particularly, Wnt7a negativity was associated with worse DFS and OS, especially in the ER-positive group. Therefore, Wnt7a could play an important role in promoting tumor progression, and its expression may be helpful in predicting outcomes and improving prognostic models.
The Wnt signaling family includes glycoproteins that are highly conserved signaling molecules important to development and tissue homeostasis. Wnt ligands bind to the Frizzled family of receptors and activate β-catenin-dependent (canonical) and β-catenin-independent (noncanonical) pathways. These signaling pathways are fundamental to control cell proliferation, cell polarity, and fate determination [5]. Wnt7a is known to induce cellular senescence via inactivation of Sphase kinase-associated protein 2, an important regulator of cellular senescence [11].
Previous studies on different types of malignancies have reported different roles of Wnt7a expression, which are either oncogenic or suppressive with respect to cancer progression. In studies where Wnt7a promoted cancer progression, Wnt7a expression was associated with aggressive behavior in renal cell carcinomas [13], mesotheliomas [14], and uterine cervical carcinomas [10]. In studies where Wnt7a suppressed cancer progression, loss of Wnt7a expression might contribute to lung cancer progression through the loss of E-cadherin via the Wnt/β-catenin signaling pathway [17]. Moreover, methylation of Wnt7a was correlated with advanced AJCC stage and high histological grade in renal cell carcinoma [13]. Given these two paradoxical actions of oncogenic function and tumor suppression, Wnt7a may play a pivotal role in the crossroads of physiological pathways in several malignancies.
In studies of hormone-dependent organ cancers such as ovarian cancer [918] and breast cancer [7], Wnt7a expression was linked to oncogenic action and had a low survival rate. However, in endometrial cancer, Wnt7a expression revealed two conflicting results [815]. In a study by Liu et al. [8], high Wnt7a expression was associated with lower DFS and OS rates. In contrast, another study showed a significant correlation between loss of Wnt7a expression and worse survival [15]. Based on experimental results [11], we hypothesized that loss of Wnt7a expression reduces tumor-suppressive cellular senescence such as apoptosis and autophagy, which increases carcinogen-induced breast tumorigenesis. However, the mechanism is limited to ER-positive breast cancer and is not related to other types of breast cancer. Here, Wnt7a expression was associated with ER positivity and a high survival rate, dissimilar to the findings of another study [7]. Especially in ERor PR-positive groups, Wnt7a expression was significantly negatively correlated with worse DFS. A significant relationship between OS and Wnt7a occurred only in the ER-positive group.
In different types of malignancy, the Wnt signaling pathway has yet to be completely explained. Wnt7a has been shown to activate both canonical and noncanonical pathways according to the cellular environment [19202122]. In the complex molecular mechanism of the Wnt signaling pathway, contrary functions such as oncogenic action and tumor suppression are complicated by mutational heterogeneity in cancer progression. Interestingly, previously published data and our results revealed that Wnt7a negativity is associated with poor clinical outcomes in ER-dependent malignancies such as endometrial and breast cancer [15]. This is because synergistic biological effects such as inhibition of cellular senescence by ER and Wnt7a facilitate cancer progression [23]. However, some exceptions to biological behaviors by Wnt7a and ER exist. Contrary to our results, other hormone-dependent cancers did not exhibit tumor-suppressive effects by Wnt7a [7918]. The functional role of Wnt7a may not apply in some parts of the senescence pathway and may depend on several environmental factors and other multiple functions in breast cancer. Furthermore, the discrepancy in correlation between Wnt7a expression and prognosis can be explained by various factors such as study design, cancer type, ethnic factors, and sample size.
Some limitations should be considered in this study. First, this cross-sectional study did not show continuous relationships over time, making it difficult to confirm a definite conclusion. Second, representative areas may not have been evaluated, because Wnt7a expression was only analyzed in one 2-mm sized core for each tumor specimen. Lastly, alteration in Wnt7a expression according to adjuvant therapeutic effects could not be evaluated, because the Wnt7a level after treatment was not investigated in fresh tumor tissue [24].
In summary, there was a significant relationship between Wnt7a expression and ER positivity in breast cancer. Moreover, Wnt7a negativity was associated with poor DFS. In the ER-positive group, Wnt7a negativity was related to worse DFS and OS, whereas in the PR-positive group, there was a relationship between Wnt7a and DFS. Therefore, Wnt7a expression may have clinical utility as a prognostic marker in hormone-dependent cancer. Relevant larger-scale studies would be useful to confirm the relationships between Wnt7a expression and prognosis in ER-positive breast cancer.
Go to :
 XML Download
XML Download