

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Breast cancer is the most common cancer and the second most common cause of cancer-related death in women [1]. Despite the successful eradication of the primary tumor, cancer can still occur in other areas of the body; therefore breast cancer metastasis is the main cause of death in breast cancer patients [2]. Invasion, which is the initial step in cancer metastasis, allows the tumor to invade into the surrounding tissue and metastasize. Thus, it is a popular target in the study of metastasis [3].
The degradation of the extracellular matrix, in which matrix metalloproteinases (MMPs) play a key role, is a critical step during cell invasion [4]. MMPs are a family of zinc endopeptidases, of which 26 members have been discovered. Among these, MMP-9 can degrade type IV collagen, which is a major structural component of basement membranes; thus, the control of MMP-9 expression is thought to be important in the regulation of metastasis [56].
Crotonis fructus (CF) is the mature fruit of Croton tiglium L., which is a leafy shrub native to Southeast Asia. CF has been used to attenuate gastrointestinal diseases such as constipation, visceral pain, and intestinal inflammation [78]. Isoguanosine from C. tiglium L. was shown to possess antitumor activity against implanted S-180 ascitic tumors in mice [9]. However, it has remained unclear whether CF extract (CFE) inhibited the invasion of breast cancer cells.
In the present study, the effects of CFE on 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced MCF-7 cell invasion were evaluated through measurements of cell invasiveness and MMP-9 activity. In addition, the signaling mechanism was investigated.
METHODS
Cells and materials
MCF-7 cells were obtained from the American Type Culture Collection (Manassas, USA). The cells were grown in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics. TPA and anti-β-actin antibodies were obtained from Sigma-Aldrich (St. Louis, USA). FBS, high-glucose DMEM, and phosphate-buffered saline were obtained from Gibco-BRL (Gaithersburg, USA).
Preparation of crotonis fructus extract
The mature fruits of C. tiglium L. were purchased from Kwangmyungdang Medicinal Herbs Co., Ltd. (Ulsan, Korea) and authenticated by professor G.S.L. The fruits were powdered and extracted with 70% aqueous ethanol under reflux for 2 hours. The solvent was removed by evaporation and the extract was freeze-dried. The yield of the final extract was 11.48% (w/w).
Cell viability assay
The effect of CFE on MCF-7 cell viability was determined by using EZ-Cytox Enhanced Cell Viability Assay Kit (DOGEN, Seoul, Korea) in accordance with the manufacturer's instructions. Briefly, 3×104 cells/well were treated with 1, 5, 10, 30, 50, or 100 µg/mL CFE for 24 hours. Thereafter, EZ-cytox was added (10 µL/well), followed by an incubation period of 30 minutes at 37℃. The absorbance at 450 nm was measured by using an ELISA reader (Sunrise™; Tecan, Männedorf, Switzerland).
Western blot analysis
The cells were incubated for 24 hours with TPA, which was followed by treatment with 10 or 30 µg/mL CFE for 1 hour. The cells were lysed with ice-cold radioimmunoprecipitation assay (RIPA) buffer (Thermo, Rockford, USA), and the protein lysates (20 µg) were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The separated proteins were transferred to polyvinylidene difluoride (PVDF) membranes using Western blotting apparatus. The membranes were blocked for 2 hours with 2% bovine serum albumin or 5% skim milk and then incubated with 1 µg/mL primary antibody overnight at 4℃. The primary antibodies used for detection were: protein kinase C (PKC)α, PKCβ, PKCδ, p38, phosphorylated p38 (p-p38), c-Jun N-terminal kinase (JNK), p-JNK, extracellular signal-regulated kinase (ERK), p-ERK, p-c-Jun (Cell Signaling Technology, Beverly, USA), sodium potassium ATPase (Na-K ATPase) (Abcam, Cambridge, UK), MMP-9, p50, p65, and proliferating cell nuclear antigen (PCNA) (Santa Cruz Biotechnology, Dallas, USA). Horseradish peroxidase conjugated IgG was used as a secondary antibody and the protein expression levels were evaluated by signal analysis using a MINI HD6 image analyzer (Uvitec, Cambridge, UK).
Gelatin zymography assay
First, the medium was collected and mixed with sample buffer. The gel for electrophoresis contained 0.1% (w/v) gelatin. The gel was washed with 2.5% Triton X-100 solution at 18℃ to 23℃ (room temperature) for 30 minutes and then incubated at 37℃ in 5 mM CaCl2, 0.02% Brij, and 50 mM Tris-HCl (pH 7.5) for 16 hours. The gel was stained using Coomassie brilliant blue solution and then photographed using an image analyzer (Fujifilm, Tokyo, Japan). Proteolysis was identified by the presence of white zones in dark blue fields. Densitometric analysis was performed by using Multi Gauge Software for Image Analysis (Fujifilm).
Quantitative real-time polymerase chain reaction
Total RNA was isolated using TRIzol® Reagent (Life Technologies, Grand Island, USA). The cDNA was synthesized from 1 µg total RNA by using the PrimeScript™ RT reagent kit (TaKaRa, Shiga, Japan). The RNA expression levels of MMP-9 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were determined using the ABI PRISM 7900 sequence detection system and SYBR® Green (Applied Biosystems, Foster City, USA). The following primers (2 µM) were used: MMP-9 (NM 004994), sense 5´-CCTGGAGACCTGAGAACCAATCT-3´, antisense 5´-CCACCCGAGTGTAACCATAGC-3´; and GAPDH (NM 002046), sense 5´-ATGGAAATCCCATCACCATCTT-3´, antisense 5´-CGCCCCACTGATTTTGG-3´.
Preparation of nuclear extract
The cells were treated with CFE in the presence or absence of TPA for 3 hours and then pelleted by centrifugation at 1,800×g for 4 minutes. The isolation of the cytoplasmic and nuclear extracts was performed using the NE-PER® Nuclear and Cytoplasmic Extraction Reagents (Thermo Scientific, Rockford, USA).
Electrophoretic mobility gel shift assay
Affinity purification of the MCF-7 nuclear protein was performed as described above. The MCF-7 nuclear protein was incubated at room temperature in 10 mM Tris-HCl, 2.5% (v/v) glycerol, 5 mM MgCl2, 50 ng/µL poly (dI-dC), and 0.05% (v/v) Nonidet P-40 binding buffer, pH 7.5. Then, 1 ng of activator protein-1 (AP-1) electrophoretic mobility gel shift assay (EMSA) probe was added (Panomics Inc., Fremont, USA). After incubation for 20 minutes at room temperature, the samples were resolved on a 4% polyacrylamide gel in 0.5× Tris-borate buffer. The proteins were electrotransferred onto a 0.45-µm nylon membrane (Hybond-N+; Amersham, UK) at 100 V for 45 minutes at 4℃ and then crosslinked to the membrane with CL-1000 ultraviolet crosslinker (UVP Inc., Upland, USA). The blots were developed by using a Chemiluminescent Nucleic Acid Detection Module Kit (Thermo Scientific).
Membrane fractionation
MCF-7 cells (5×107) were pretreated with 30 µg/mL CFE for 1 hour and then incubated with TPA for 1 hour at 37℃. The membrane proteins were prepared from cells using the MEM-PER™ plus kit (Thermo Scientific).
Invasion assay
The invasion assay was conducted in 24-well chamber dishes (8-µm pore size) coated with 20 µL of Matrigel that was rehydrated in 0.5 mL DMEM for 30 minutes prior to the experiments. Cells (2×105) and chemoattractant (conditioned medium; 0.5 mL) were added to the upper and lower chamber of the dishes, respectively. After incubation, the invading cells were fixed and stained with a Toluidine blue solution. The analyzed data were presented as the mean±standard error from three individual experiments performed in triplicate.
RESULTS
Effect of crotonis fructus extract on MCF-7 cell viability
To evaluate the cytotoxicity of CFE on MCF-7 cells, the viability of untreated and CFE-treated cells was compared. At all tested concentrations, CFE treatment for 24 hours did not result in any significant changes in cell viability. CFE also did not affect MCF-7 cell proliferation at 100 µg/mL for up to 72 hours (Figure 1A and Supplementary Figure 1A, available online). Thus, the optimal nontoxic concentrations (10 and 30 µg/mL) of CFE were used for all subsequent experiments.
Effect of crotonis fructus extract on TPA-induced MMP-9 expression/secretion in MCF-7 cells
To examine the effect of CFE on TPA-induced MMP-9 expression, MCF-7 cells were analyzed by Western blot analysis, real-time quantitative polymerase chain reaction (RT-qPCR), and gelatin zymography. Western blot analysis revealed that CFE treatment blocked the upregulation of TPA-induced MMP-9 protein expression in MCF-7 cells. Zymography was performed to determine the effects of CFE on TPA-induced MMP-9 secretion and showed that TPA increased MMP-9 secretion in MCF-7 cells. However, CFE significantly diminished TPA-induced MMP-9 secretion (Figure 1B). RT-qPCR revealed that TPA induced an increase in MMP-9 expression levels in MCF-7 cells, whereas CFE blocked TPA-induced MMP-9 upregulation in a dose-dependent manner (Figure 1C). These results indicated that CFE exerted potent inhibitory effects against the TPA-induced upregulation of MMP-9 expression in MCF-7 cells.
Effect of crotonis fructus extract on TPA-induced PKC activation in MCF-7 cells
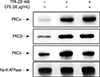
To determine the effects of CFE on TPA-induced PKC activation, we analyzed the PKC isotype levels in the membrane fraction. CFE inhibited the membrane localization of PKCδ induced by TPA (Figure 2), which suggested that CFE inhibited TPA-induced MMP-9 expression via the suppression of PKCδ activation in MCF-7 cells.
Effect of crotonis fructus extract on TPA-induced MAPK activation in MCF-7 cells
To further understand the signaling pathway involved in the inhibitory action of CFE on TPA-induced MMP-9 activation, we used Western blotting to evaluate the effect of CFE on the TPA-induced activation of mitogen-activated protein kinase (MAPK). The treatment with CFE showed inhibitory effects on p38 and JNK, but not on ERK (Figure 3A). These results suggested that the p38 and JNK pathways were involved in the suppression of TPA-induced MMP-9 expression by CFE.
Effect of crotonis fructus extract on TPA-induced activation of NF-κB and AP-1 in MCF-7 cells
It has been reported that the MMP-9 promoter has nuclear factor-κB (NF-κB) and AP-1 binding sites; these are known to be involved in MMP-9 gene induction by TPA [1011]. To clarify the transcription mechanism by which CFE inhibited MMP-9 expression, the effect of CFE on the TPA-induced activation of NF-κB and AP-1 was evaluated using Western blotting. As shown in Figure 3B, TPA upregulated the expression of p65 and p50 (NF-κB subunits) and p-c-Jun (AP-1 subunit) in the nuclear fraction. The pretreatment of cells with CFE resulted in the inhibition of TPA-induced p-c-Jun expression, but not that of NF-κB activation. Additionally, the DNA binding activity of AP-1 was confirmed. The treatment with CFE suppressed TPA-induced AP-1 DNA binding activity (Figure 3C). These results indicated that CFE specifically blocked AP-1 activation in MCF-7 cells.
Effect of crotonis fructus extract on TPA-induced cell invasion in vitro
The upregulation of MMP expression is known to contribute significantly to cancer invasion [1213]. In this study, we investigated the inhibitory effects of CFE on the invasive ability of MCF-7 cells induced by TPA by performing a Matrigel invasion assay. The cells pretreated with 20 nM TPA were subsequently incubated with CFE for 24 hours. TPA-induced MCF-7 cell invasion was suppressed by CFE treatment (Figure 4). Additionally, CFE slightly inhibited the migration and invasion of MDA-MB-231 cells, a highly metastatic cell line, at nontoxic concentration (Supplementary Figure 2, available online).
DISCUSSION
The main components of CF, the mature fruit of C. tiglium L., are croton oils, which are traditionally used as a laxative or purgative. This oil contains a toxic resin, which is a highly toxic substance that must be treated with great caution. Thus, croton oil can lead to initial skin sloughing, inflammation, and pain [14]. For that reason, the croton oils in CF are removed. Although there are limited studies on the effects of CFE, the effectiveness of CFE for the therapy of peptic ulcers and for the stimulation of lipolysis has been reported [1516].
MMP-9 is a well-known proteinase that disrupts the extracellular matrix, which induces the invasion of cancer cells; thus, it has been widely studied as a major target of cancer metastasis. In the present study, we found that CFE treatment suppressed the TPA-induced protein, mRNA expression, and secretion of MMP-9 (Figure 1). These results were the first to demonstrate the effect of CFE on MMP-9.
As described above, MMP-9 expression was closely correlated with cancer cell invasion. The present findings showed that CFE blocked TPA-induced MCF-7 cell invasion (Figure 4). Additionally, CFE inhibited TPA-induced MCF-7 migration (Supplementary Figure 1B, available online). As this was the first study to demonstrate the inhibitory activity of CFE on cell invasion, the mechanism of action was also evaluated. The activation of the three major MAPK families, JNK, ERK, and p38 kinase, is required for MMP-9 induction [1718]. The MMP-9 gene promoter contains both NF-κB and AP-1 binding sites. AP-1, composed of Jun and Fos subunits, can be activated by p-JNK [1920]. As shown in Figures 2 and 3, CFE inhibited the TPA-induced phosphorylation of p38, JNK, and c-Jun. These results showed that CFE suppressed TPA-induced MMP-9 expression through the MAPK/AP-1 pathway in MCF-7 cells.
It was previously reported that PKC isoforms contributed to the TPA-induced invasion and migration of MCF-7 cells, suggesting that the TPA-induced upregulation of MMP-9 expression and cell invasion involved the activation of PKCδ and PKCα [21]. In this study, the translocation of PKCδ into the cell membrane was slightly inhibited by CFE treatment (Figure 2A). These results suggested that CFE possessed the ability to regulate the activation of TPA-responsive PKC isozymes, especially PKCδ, in MCF-7 cells.
This study demonstrated that CFE exhibited anti-invasive activity in TPA-induced MCF-7 cells and suppressed TPA-induced MMP-9 expression via the activation of the PKCδ/p38/JNK/AP-1 pathway. Therefore, the anti-invasive effect of CFE on breast cancer cells at nontoxic concentrations indicated its potential as a chemopreventive agent against breast cancer.


 XML Download
XML Download