

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Breast cancer is the most common malignancy in women worldwide [1]. Although breast cancer therapy has significantly improved during the last decade, the mortality rate associated with this disease remains quite high. Recent advances in epigenetics have demonstrated that histone modifications, especially histone acetylation and deacetylation, may lead to improved treatment strategies in breast cancer. Histone acetyl-ation and deacetylation are predominantly modulated by histone acetyltransferases (HATs) and histone deacetylases (HDACs), respectively [2]. HDACs remove acetyl groups from histone proteins, and have been suggested to exert a pro-oncogenic effect, whereas HATs have the opposite effect [345]. Therefore, cancer progression is widely perceived to be dependent on the balance between HAT and HDAC activity. In this context, the deregulation of HAT has been shown to be associated with cancer development [6]. Accordingly, exploring the underlying mechanisms or factors that can influence histone acetylation may lead to the identification of new and promising targets for breast cancer research.
Transformation/transcription domain-associated protein (TRRAP) is present in the HAT complex and assists with recruitment of the HAT complex to chromatin during gene transcription [78]. Studies have shown that TRRAP is not only a common component of HAT complexes in yeast, but also in mammalian cells [9]. Thus, it is reasonable to assume that TRRAP has the ability to modulate the function of the HAT complex.
TRRAP was originally identified as an interacting partner of the c-Myc protein [10]. It has been suggested that TRRAP is an adaptor protein homologous to phosphatidylinositol 3-kinase-related kinases (PIKKs) but without intrinsic kinase activity. It has been shown to contribute to various biological functions such as cell cycle progression, oncogenic transformation via c-Myc and E2F, chromatin remodeling, and embryonic development. TRRAP has also been shown to be an important component of the BRCA1 (breast cancer 1, early onset) genome surveillance repair complex and impairment of BRCA1 function predisposes women to early onset of breast and ovarian cancer [11]. Recent studies have demonstrated that overexpression of TRRAP in gliomas is associated with promotion of stem cell characteristics, and TRRAP is usually elevated in gliomas [12].
Since TRRAP is an important component of HATs, which are important in the development of cancer, and can enhance stem cell-like characteristics as well as regulate BRCA1 gene function, we hypothesized that TRRAP might play an important role in cancer. Therefore, this study investigated the association between TRRAP and breast cancer, and its clinicopathological features.
METHODS
Patients and clinical samples
We obtained 470 patient tissue samples of confirmed histology of breast cancer, and 244 normal tissue samples, from the Department of Pathology at Affiliated Tumor Hospital of Harbin Medical University. Informed consent was obtained from all patients. The Institutional Review Board of Affiliated Tumor Hospital of Harbin Medical University approved this study (KY 2013–29). Tumor and normal breast tissues were examined diagnostically by two pathologists. All patients had invasive breast cancer, and all samples were collected before any radiotherapy or chemotherapy was applied. The tumor size at the largest diameter of the invasive carcinoma was measured in millimeters by the pathologists. Normal breast tissues were collected at least 5 cm away from the tumor tissues and were further confirmed to be cancer free. The tissue sections were prepared from formalin-fixed and paraffin-embedded blocks.
All selected patients had complete medical records from 2006 onwards. Each individual sample was analyzed by immunohistochemistry for estrogen receptor (ER), progesterone receptor (PR), human epidermal growth factor receptor 2 (HER2), Ki-67, and p53 expression. Immunohistochemical staining for ER and PR proteins was performed using a conventional detection method. Greater than 10% nuclear staining in the invasive component of tumor cells represented a positive result [13]. In addition, individual tumor samples with ≥14% Ki-67 positive staining were considered highly proliferative [14].
The intensity of anti-HER2 staining in all samples was semiquantitatively analyzed and graded as 0 to 3. Individual samples with a grade of 0 were considered negative while samples with grade 1, 2, or 3 were regarded as positive.
Examinations of all the patients were performed every 4 to 6 months for the first 5 years and every 12 months thereafter. Patients were followed regularly for a minimum of 5 years or until death or the study closing date (December 30, 2012) at the Affiliated Tumor Hospital of Harbin Medical University. Overall survival was assessed to determine prognosis.
Tissue microarray generation and immunohistochemical staining
Breast cancer and normal tissue microarrays were created by punching a hole in receptive paraffin block using a thin-walled needle with an inner diameter of 2 mm, to acquire tissue cores from the tissue block. After construction of the array block, all tissue blocks were cut with a microtome to a thickness of 4 µm.
The tissue sections were dried at 70℃ for 3 hours. This was followed by deparaffinization and hydration. Subsequently the sections were washed with phosphate-buffered saline (PBS; 3×3 minutes). The washed sections were treated with 3% H2O2 in the dark for 5 to 20 minutes and again washed with distilled water. The sections were further washed with PBS (3×5 minutes) and antigen retrieval was performed in citrate buffer (pH 6.0). This was followed by incubation of each section with 300 to 500 µL TRRAP antibody (1:250; Abnova Inc., Taipei, Taiwan) at 4℃ overnight. The next morning, after washing with PBS (3×5 minutes), each section was further incubated with 300 to 500 µL of secondary antibody (1:200; Abcam, Cambridge, England) at room temperature for 30 minutes. After washing with PBS (3×5 minutes) again, each section was incubated with 300 to 500 µL of 3,3'-diaminobenzidine (DAB) working solution at room temperature for 3 to 10 minutes, and then finally the sections were washed with distilled water.
Expression levels were assessed based on the staining intensity and distribution. The staining intensity was graded as 0, no staining; 1, weak staining, light yellow; 2, moderate staining, yellow brown; or 3, strong staining, brown. The percentage of reactivity was scored as 0 (no positive tumor cells); 1 (fewer than 10% positive tumor cells); 2 (10%–50% positive tumor cells); and 3 (more than 50% positive tumor cells) [15]. Based on these criteria, the overall expression level was scored by multiplying the intensity and reactivity values. Scores <4 reflected low expression, while all scores >4 were indicated high expression.
Inspection of all samples by pathologists suggested that >80% of the cells in each section were cancer cells. Finally, TRRAP protein expression was assessed by evaluating the proportion and intensity of staining in a series of 10 randomly selected high-power fields, which were considered representative of the average in a ×400 magnification field. Two investigators without knowledge of the clinicopathological findings scored the staining pattern of each sample independently.
Statistical analysis
The chi-square test was performed to compare the data from different groups. A two-sided p-value of <0.05 was considered statistically significant. Data were analyzed using the SPSS software version 17.0 (SPSS Inc., Chicago, USA). Overall survival was estimated using the Kaplan-Meier method.
RESULTS
TRRAP expression in breast carcinoma tissues
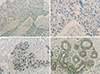
We compared the protein expression of TRRAP in breast carcinomas to that of corresponding normal breast tissues. TRRAP expression levels in invasive ductal breast carcinoma samples (Figure 1A, B) were lower than that in matched normal breast tissue samples (Figure 1C, D). A large number of samples from both categories was quantified for TRRAP expression (Table 1), and the data showed a significantly lower TRRAP expression in invasive ductal breast carcinomas than in normal tissues (p<0.001).
Association between TRRAP expression and clinicopathological features
We analyzed the association between TRRAP expression and a series of clinicopathological characteristics, including patient and tumor characteristics (Table 2). Among the breast cancer patients diagnosed with invasive ductal carcinoma, 298 patients (63.4%) tested negative for TRRAP, whereas 172 (36.6%) tested positive (Table 1). There was significantly greater nodal involvement in patient samples with no TRRAP expression (42.6%) than in those positive for TRRAP (27.3%). The mean tumor size in TRRAP negative samples was 2.83±1.60, which was significantly larger than the size observed in samples positive for TRRAP expression (2.48±1.44).
An investigation of the association between TRRAP expression and differentiation grades also revealed some significant differences. Tumors that were negative for TRRAP expression were more likely to be less differentiated than those positive for TRRAP. Specifically in tumors with no TRRAP expression, 16.4%, 44.6%, and 38.9% of the tumors were well, moderately, and poorly differentiated, respectively. However, in tumors positive for TRAAP, the corresponding percentages were 22.7%, 55.2%, and 22.1%, respectively.
The rate of positive p53 expression was significantly lower in tumors with negative TRRAP expression (77.9%) than in those with positive TRRAP expression (89.0%) (Table 2). However, levels of ER, PR, HER2, and Ki-67 were similar between tumors regardless of TRRAP expression level.
The correlation between TRRAP expression and disease prognosis
We assessed the potential clinical significance of TRRAP expression in breast cancer prognosis by performing univariate and multivariate analyses (Table 3). Univariate analysis demonstrated that TRRAP expression and PR were independent protective factors for breast cancer prognosis, while Ki-67, tumor size, and nodal involvement were independent risk factors. Similarly, the multivariate Cox regression model also revealed that TRRAP expression, Ki-67, and nodal involvement were independent prognostic markers. Moreover, the Kaplan-Meier survival curve analysis suggested that lack of TRRAP expression was associated with shorter survival times (Figure 2).
DISCUSSION
Despite improvements in cancer therapeutics, a large number of patients diagnosed with invasive breast carcinomas will eventually die from this disease. Therefore, further identification of effective molecular targets and biomarkers for tumor classification is urgently required. Therefore, in this study, we investigated the role of the TRRAP protein in invasive ductal breast carcinomas, and analyzed its association with different clinicopathological features. Our data revealed that TRRAP expression levels in breast carcinoma tissue were lower than in normal patient-matched tissues. Moreover, in these breast cancer patients, the expression levels of TRRAP were also associated with tumor size, grade, nodal involvement, and p53 expression. There was no significant association between TRRAP expression and specific breast cancer subtypes. In add-ition, we also found that negative TRRAP expression was associated with shorter survival times. Thus, our analysis suggests that TRRAP expression may be negatively involved in breast tumorigenesis.
The molecular mechanisms regarding how TRRAP influences the development of breast cancer has been unclear. As shown in Figure 3, we explored the different interaction partners (either direct or indirect) of TRRAP using the Ingenuity Pathway Analysis (IPA) software (QIAGEN, Duesseldorf, Germany). The analysis speculated that TRRAP can interact and may regulate the function of many genes, such as MYC, BRCA1, CTNNB1, KAT2A (K[lysine] acetyl transferase 2A), KAT2B, E2F4, and p53.
Among these genes, as a confirmation of the IPA analysis, we observed that in our study, p53 expression was significantly lower in the tumors of patients that tested negative for TRRAP expression and vice-versa. The p53 protein has been shown to be an important tumor suppressor gene and acts as a key transcription factor in cellular stress response pathways [1617]. An earlier study showed that p53 can be acetylated by HATs and deacetylated by HDACs [18]. Enhancement of p53 acetylation leads to protein stabilization and activation [19]. The balance between p53 acetylation and deacetylation is often disrupted in diseases such as cancer [20]. Another study revealed that pharmacologic activation of HATs promotes cancer cell apoptosis through direct hyperacetylation of p53 [21]. In breast cancer cells, acetylation of p53 has been shown to induce cell death [22]. Based on all these published findings, and the present observations, it would be reasonable to deduce that loss of TRRAP expression can promote deacetyl-ation of p53 by inactivating HATs, eventually leading to the development of breast cancer.
However, other TRRAP interacting partners such as KAT2A and KAT2B are histone acetyl transferases and primarily function as transcriptional activators [23]. E2F2 and E2F4 belong to the E2F family and are involved in control of the cell cycle and act as tumor suppressor proteins [24]. BRCA1 impairment predisposes the onset of ovarian and breast cancer, and has been shown to interact with TRRAP, which might be involved in BRCA1 gene regulation [11]. Thus, these interacting partners further support the hypothesis that TRRAP is a tumor suppressor in cancer. However, additional studies are warranted to explore the specific function of TRRAP in the development of breast cancer.
The breast cancer field currently analyzes everything through a lens of specific molecular subtypes, especially in the clinic. The different molecular subtypes are based on the expression levels of ER, PR, and HER2 protein markers. Breast cancer that is negative for all three markers (i.e., triple-negative breast cancer) has attracted considerable attention due to its aggressive biological behavior and poor clinical outcome [25]. In this context, we explored the possibility of an association between TRRAP protein expression and the different breast cancer subtypes and found no significant association.
Finally, we also explored the association between TRRAP and various other clinical outcomes. The Kaplan-Meier survival curve data showed that positive TRRAP expression was associated with improved survival. This observation suggests that TRRAP may suppress or inhibit the proliferation of breast cancer cells. After adjusting for the influence of other confounders, multivariate Cox proportional hazard model analysis showed that positive TRRAP expression could reduce the risk of death by 49.1% as opposed to negative TRRAP expression. This information further confirmed that lack of TRRAP expression was associated with poor breast cancer prognosis.
In conclusion, our results indicate that the presence of TRRAP protein negatively correlates with breast carcinoma progression and thus seems to act as a tumor suppressor. To the best of our knowledge, this is the first detailed study that simultaneously analyzed TRRAP expression in a large number of normal and breast cancer tissues, and investigated a correlation to clinical outcomes, such as overall survival in breast cancer patients. Further understanding of the functional and molecular properties of TRRAP will aid in providing greater insight into its role in breast tumorigenesis.





 XML Download
XML Download