

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The S100 protein family consists of over 20 family members of low molecular weight calcium binding proteins [1]. These proteins share common amino acid sequence domains and a functional EF-hand motif that plays a key role in calcium binding through a helix-loop-helix topology [23].
Hornerin is a member of the S100-fused protein family. It was first identified in the mouse embryo epidermis. It was also detected in the skin, tongue, esophagus, and proximal stomach of adult mouse tissues. Hornerin has a calcium binding EF-hand structure at the N-terminus followed by a spacer sequence and a large repetitive domain [4]. The hornerin protein is similar to profilaggrin (which is involved in cornification of keratinocytes) in its structural features, expression profiles, extensive posttranslational proteolytic processing, and tissue localization [56]. Hornerin has been thought to be involved in pre- and post-natal mammary development in breast tissue [7]. Although the physiological role of S100 proteins has not been clearly identified, recent experimental data have suggested that they are involved in physiological and pathological processes such as gene transcription, inflammatory and immune responses, regulation of protein phosphorylation, transcription factors, antimicrobial responses, calcium homeostasis, dynamics of cytoskeleton constituents and, cell proliferation, differentiation, and death [8]. Differential expression of the S100 family members in certain cancers has been reported [1]. Experiments focusing on their role in mammary carcinogenesis have demonstrated that the expression levels of some S100 family proteins in basal-type breast cancers are higher than that in nonbasal types breast cancers [89]. Upregulated expression of hornerin in less aggressive breast cancers has been reported [8910]. S100A8 and S100A9 are known to be associated with high grade and basal-type breast cancer [8910]. It is generally accepted that breast cancers arise by a multistep process in which normal epithelial cells transform into invasive cancer via atypical ductal hyperplasia and in situ carcinoma, though it is not necessarily a linear process [1112]. However, the roles of hornerin, S100A8, and S100A9 in the progression of mammary carcinogenesis have not been fully evaluated. Therefore, the objective of this study was to determine their expression levels in ductal carcinoma in situ (DCIS), invasive ductal carcinoma (IDC), and metastatic carcinoma in the same patient to clarify their roles in cancer progression.
METHODS
Tissue specimens
A total of 94 cases of surgically resected IDC at Korea University Guro Hospital during 2007 to 2011 were included in this study with approval from the Institutional Review Board of the hospital (IRB number: KUGH 12149). All subjects had invasive carcinoma, adjacent DCIS component, and lymph node metastasis. Hematoxylin and eosin-stained slides for each case were reviewed for tumor subtype, histologic grade, nuclear grade, and lymph node status. The medical records of all subjects were reviewed. Nottingham's histologic grade and nuclear pleomorphism score were analyzed in this study.
Clinicopathologic information was obtained by reviewing medical records, pathology reports, and hematoxylin and eosin-stained sections. The following histopathologic variables were determined in IDCs: tumor subtype, pT stage, pN stage, Nottingham combined histologic grade [13], estrogen receptor (ER), and human epidermal growth factor receptor 2 (HER2). Tissue microarrays (TMAs) were constructed using two representative cores (2.0 mm in diameter) of primary IDCs, adjacent DCISs, or metastatic carcinomas from the same case.
Immunohistochemical analysis and silver in situ hybridization
Immunohistochemical (IHC) analyses of hornerin, S100A8, and S100A9 were performed using the Bond-Max system (Leica Biosystems, Wetzlar, Germany). Antigens were retrieved according to the Bond Max ER1 antigen retrieval protocol. Antibodies used in this study included those against hornerin (rabbit polyclonal anti-human antibody, dilution 1/200; Novus Biologicals, Littleton, USA), S100A8 (mouse anti-human antibody, 1/800; Lifespan Bioscience, Seattle, USA), and S100A9 (goat polyclonal anti-human antibody, 1/400; Santa Cruz Biotechnology, Santa Cruz, USA).
The percentage of tumor cells exhibiting intense staining for hornerin, S100A8, and S100A9 were determined in 10 high-power fields. Cases were considered positive when more than 10.0% of tumor cells were stained or negative when 10.0% or less were stained [14]. As tumor heterogeneity can exist, any expression of protein at more than 10.0% in two TMA cores was interpreted as positive.
IHC analyses of ERs (Ventana Medical Systems, Tucson, USA) and HER2s (Ventana Medical Systems) were performed using the Ventana BenchMark automatic staining system (Ventana Medical Systems). Cancer cells with ER staining in the nucleus were considered immunoreactive and scored. The evaluation of hormone receptor expression was based on the Allred scoring method and the American Society of Clinical Oncology/College of American Pathologists (ASCO/CAP) guidelines [15]. For HER2, membranous staining was also evaluated according to the guidelines of ASCO/CAP. Cases with a score of 3 were considered HER2-positive, whereas those with a score of 2 were evaluated for HER2 gene amplification according to ASCO/CAP guidelines. Silver in situ hybridization (SISH) was performed with a Ventana BenchMark automated instrument (Ventana Medical Systems) according to the manufacturer's protocols using INFORM HER2DNA probe (Ventana Medical Systems) or chromosome 17 probes (Ventana Medical Systems). These probes were labeled with dinitrophenol (DNP) and visualized using rabbit anti-DNP primary antibody and the ultraView SISH Detection Kit. Briefly, the HER2 DNA probe was denatured at 95℃ for 4 minutes and hybridized at 52℃ for 2 hours. The chromosome 17 probe was denatured at 95℃ for 4 minutes and hybridized at 44℃ for 2 hours. The final reaction was driven by the sequential addition of silver acetate, hydroquinone, and hydrogen peroxidase to the peroxidase-conjugated goat anti-rabbit antibody in the detection kit to produce a silver precipitate, which was deposited into the HER2 genes. Red centromeric signals in chromosome 17 were seen as red dots. For SISH test, we defined HER2 positivity as HER2 gene amplification by SISH with a gene copy ratio of HER2:chromosome 17 centromere ≥2.0 as described previously [16].
Statistical analysis
Statistical analyses were performed using the SPSS version 12.0 for Windows (SPSS Inc., Chicago, USA). Pearson chisquare test (or Fisher exact test when appropriate) was used to compare the binary categories of hornerin, S100A8, and S100A9 expression between groups. Paired t-tests were performed to determine whether there were significant differences between the mean percentages of hornerin, S100A8, and S100A9 expression in DCIS, IDC, and metastatic carcinoma in lymph node (MCN). McNemar test was used to assess the correlation between S100A8 and S100A9. Data were considered statistically significant when the p-value was less than 0.050.
RESULTS
Demographic and pathologic data (clinicopathologic data)
From the 94 cases, the data that we required were available for 70 cases. The median age of the 70 patients was 51 years (range, 33–83 years). Clinicopathologic features are shown in Table 1. According to the seventh edition of the American Joint Committee on Cancer Staging Manual [17], the primary tumor pT1 occurred in 40.0%, pT2 in 55.7%, pT3 in 4.3%, pN1 in 55.7%, pN2 in 34.3%, pN3 in 10.0%, TNM stage 0/IA/IB in 0.0%, IIA in 22.9%, IIB in 31.4%, IIIA in 35.7%, IIIB in 0.0%, and IIIC in 10.0% of cases. Based on the Nottingham histologic grade, the distribution of grades was as follows: grade I in 21.4%, grade II in 50.0%, and grade III in 28.6% of cases. Based on the nuclear grade of the primary tumor, the distribution of grades was as follows: grade I in 2.9%, grade II in 70.0%, and grade III in 27.1% of cases.
Expression of hornerin, S100A8, and S100A9 in breast cancer
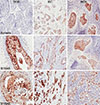
Hornerin, S100A8, and S100A9 were easily detectable in the cytoplasm and focally positive in the nucleus of tumors (Figure 1). Hornerin was positive in 15 of 70 IDCs (21.4%). S100A8 was positive in 22 of 70 IDCs (31.4%), and S100A9 was positive in 39 of 70 IDCs (55.7%).
Hornerin positivity in IDC was significantly higher in ERnegative (p=0.003) and HER2-positive groups (p=0.002). However, hornerin expression in IDC did not show any significant relationship with stages or grades. Expression of S100A8 was associated with a higher pT stage (p=0.017); however, it was not correlated with other prognostic factors. There was no significant relationship between S100A9 expression and prognostic markers (Table 2). A significant (p < 0.001) correlation was found between S100A9 and S100A8 expression (Table 3). The number of S100A9-positive IDC cases was higher than that of S100A8-positive IDC cases. Interestingly, S100A8 was not expressed in the absence of S100A9 expression. Hornerin showed no significant (p > 0.050) correlation with S100A8 or S100A9 expression (data not shown).
Hornerin expression is increased during breast cancer progression
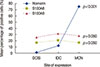
Next, we investigated whether changes in the expression of hornerin, S100A8, and S100A9 were correlated with cancer progression. The mean percentages of hornerin-positive tumor cells in DCIS, IDC, and MCN were 1.0%±3.3% (mean±standard deviation), 12.0%±24.0%, and 75.3%± 27.6%, respectively. Hornerin expression was significantly (p<0.001) higher in the MCN group compared to that of the DCIS or IDC group (Figure 2). Expression of hornerin appeared to increase in a stepwise manner (DCIS<IDC< MCN). The mean levels of S100A8 and S100A9 expression in DCIS, IDC, and MCN were not significantly (p>0.050) different (Figure 2).
DISCUSSION
Previous studies on "fused gene"-type cornified envelop precursor proteins have mainly focused on their physiological and pathologic roles in keratinization [461819]. However, recent studies have reported that "fused gene"-type S100 proteins may also be involved in the pathogenesis of neoplasia [1020]. The objective of this study was to determine the expression levels of hornerin, S100A8, and S100A9 in DCIS, IDC, and metastatic carcinoma in the same patient to clarify their roles in cancer progression. Our results revealed a positive correlation between hornerin expression and ER-negativity with HER2-amplification, in agreement with the results of an in vitro study that used the MCF10A cancer progression model to show that hornerin mRNA expression increased as tumorigenicity progressed [10]. However, in this study, we confirmed our results with IHC staining using formalin fixed paraffin embedded tissue and found that hornerin expression was correlated with the lobular carcinoma or node-negative group. To demonstrate the changes in expression, we specially constructed TMAs from DCIS, IDC, and metastatic lesion in lymph nodes of the same patients. IHC analyses of TMAs showed that hornerin expression differed in breast tissue according to the progression stages in the same patient, including preinvasive in situ, invasive, and metastatic. In addition, the expression of hornerin dramatically increased in a stepwise manner as breast cancer progressed from DCIS to MCN. These findings strongly suggest that hornerin is involved in breast cancer progression, specifically in the transformation from preinvasive carcinoma to invasive carcinoma. Furthermore, it was remarkable to find that the expression levels of hornerin in IDC were much higher than those in DCIS. Recently, it has been proposed that exosomes are involved in the formation of organ-specific metastatic niches [21]. Therefore, as an exosome protein, hornerin may play an essential role in the invasive process of mammary cancer progression, suggesting that it might be used as a potential marker to predict recurrence or potential metastasis. In this study, a survival analysis for hornerin in breast cancer was not performed. Therefore, further study with survival data is necessary.
S100 proteins are low molecular weight proteins ranging in size from 9 to 13 kDa. Most S100 proteins can form heteroand homo-dimers that might be essential for the generation of their active form [22]. S100A8 and S100A9 proteins usually form a heterodimer called calprotectin that is present in high levels in endothelium, macrophages, and neutrophils [232425]. Our study showed a significant correlation between S100A8 and S100A9 expression. This finding is in accordance with a previous study [8]. Since S100A8 and S100A9 proteins have been known to exist as heterodimeric molecules [26], it would not be surprising if their expression was coordinately controlled. In the present study, a positive correlation between S100A8 expression and higher pT stage was demonstrated. However, no association between S100A9 and clinicopathologic parameters with prognostic impact was observed. Some studies have indicated that overexpression of S100A8 and S100A9 is associated with poor prognosis of IDC and lung adenocarcinoma [927]. Another study has suggested an association between hornerin expression and ER-negative or high grade tumors [8].
There are some differences between our study and another study with regard to how protein expression was interpreted [10]. We interpreted it to be positive when we observed more than 10.0% of protein expressed in two TMA cores because any resultant subclone could have a specific biologic behavior due to evolutionary changes among cancer cells. As we used TMA with a unique design, including in situ, invasive, and metastatic lesions from each patient, we only enrolled a small number of cases in this study. Furthermore, we only evaluated 70 of the 94 cases recruited for this study because not all the cores fulfilled the required data.
In this study, the expression of hornerin steadily increased during breast cancer progression from preinvasive lesions to metastatic carcinoma. In addition, hornerin was expressed at higher levels in ER-negative but HER2-positive groups. These findings suggest that hornerin may be involved in mammary cancer progression. Therefore, it might be used as potential marker for poor prognosis in breast cancer management. Recently, it has been proposed that exosome proteins, such as S-100 family members, are involved in the formation premetastatic niches at metastatic sites [21]. The relationship between exosomes and S100 family members in cancer progression is starting to emerge. To clarify the roles of S-100 family members including hornerin in cancer progression, further studies with a large number of cases along with survival data are needed.



 XML Download
XML Download