

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cancer stem cells (CSCs) have recently been identified in various malignant tumors ranging from leukemia to solid tumors such as breast cancer. Evidence indicates that CSCs play an important role in the pathogenesis, invasion, and metastasis of malignancies [1]. CSCs represent only a small fraction of a tumor. However, unlike most cancer cells, they possess self-renewal and differentiation capabilities, which lead to high tumorigenesis and resistance to standard chemotherapeutic agents. Breast cancer cells with the CD44+/CD24- cell surface marker expression profile have been proposed to be CSCs [2]. Evidence has demonstrated that breast cancer cells with CD44+/CD24- have strong tumor initiation capabilities. Breast cancer cells with CD44+/CD24- are resistant to chemotherapy [3] and radiotherapy [4] because they are mostly quiescent, as they are arrested at the G0/G1 phase in mitosis. Inducing the breast cancer cells differentiation with CD44+/CD24- from G0/G1 to S may make these cells easy to destroy with chemotherapy.
We try to provide a strategy to induce breast cells differentiation with CD44+/CD24- from G0/G1 to S. Basic fibroblast growth factor (bFGF) is a widely active and potent mutagen. We propose the hypothesis that bFGF can be used to induce breast cancer cells with CD44+/CD24- to differentiate into dividing cancer cells.
Molecular markers have been used to identify breast CSCs, and the CD44+/CD24-/low phenotype has stem cell properties [5]. A single breast cancer cell marked by CD44+/CD24- can reconstitute into a breast tumor [6]. Breast cancer cells with CD44+/CD24- also have highly invasive properties and express higher levels of pro-invasive genes [7].
In this study, we report that bFGF regulates progression of the G1/S phase, and a potential method to induce breast cancer
cells with CD44+/CD24- to distinguish cancer cells from MCF-7 cells.
Go to : 
METHODS
Cell culture
The MCF-7 human breast cancer cell line was obtained from the Institute of Histoembryology, Binzhou Medical University. The cells were maintained in RPMI 1640 (Boster, Wuhan, China) containing 10% fetal bovine serum and an antibiotic (Figure 1A).
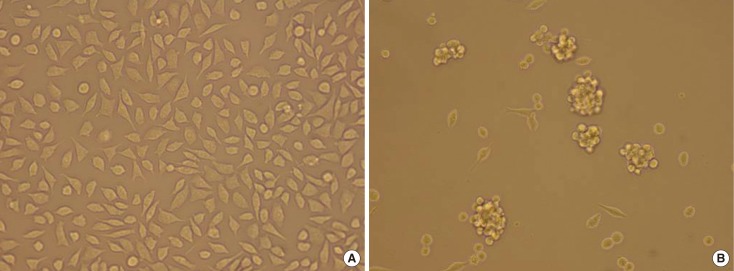 | Figure 1(A) MCF-7 cells culture. Some of the MCF-7 cells have fusiform shape, and some have polygonal shape. they array orderly in general (×20). (B) MCF-7 mammosphere culture. MCF-7 cells cultured in the medium that was supplemented with 2% B27, 5 µg/mL bovine insulin, 10 ng/mL basic fibroblast growth factor 2, and 20 ng/mL epidermal growth factor at a density of 2,000 cells/mL generate mammosphere (×20).
|
Mammosphere culture
Cells in the supernatant of 2-day-old cultures were collected by centrifugation, for 5 minutes at 300×g, washed in Hanks' buffered salt solution, and re-suspended in phenol red-free DMEM-F12 (Boster) supplemented with 2% B27 (Gibco, Grand Island, USA), 5 µg/mL bovine insulin (Boster), 10 ng/mL bFGF (PeproTech, Rocky Hill, USA), and 20 ng/mL epidermal growth factor (PeproTech) at a density of 2,000 cells/mL to obtain cancer-initiating cells and propagate them as mammospheres. Growth factors were added to the mammosphere cultures (Figure 1B) every 3 days.
Flow cytometry
Cells were washed once with phosphate-buffered saline (PBS) and then harvested with 0.05% trypsin/0.025% EDTA. Detached cells were washed with PBS containing 1% fetal calf serum and 1% penicillin/streptomycin (wash buffer), and then re-suspended in wash buffer (106 cells/100 µL). Fluorochrome-conjugated monoclonal antibodies fluorescein isothiocyanate (FITC) anti-human CD44 and phycoerythrin (PE) anti-human CD24, or their respective isotype controls, were added to the cell suspension at concentrations recommended by the manufacturer, and incubated at 4℃ in the dark for 30 to 40 minutes. The labeled cells were washed in wash buffer, fixed in PBS containing 1% paraformaldehyde, and analyzed on a fluorescence activating cell sorter (FACS)-Vantage flow cytometer (BD Biosciences, San Diego, USA). The cells were sorted into CD44+/CD24- and CD44+/CD24+ subpopulation, according to their surface markers (Figure 2). Cells were routinely sorted twice and re-analyzed for purity, which was typically >95%.
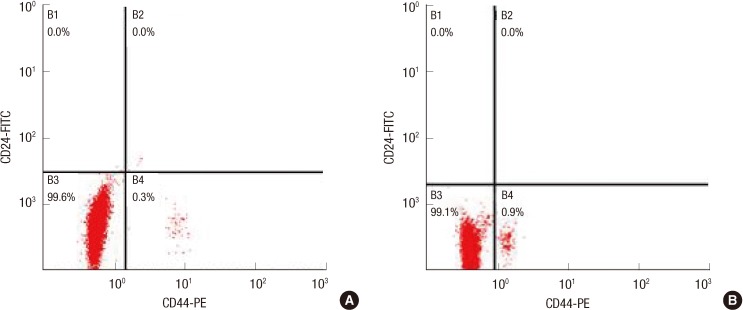 | Figure 2(A) CD44+/CD24- cells subpopulation of unselected MCF-7. Flow cytometric analysis of MCF-7 cells labeled with CD24-FITC and CD44-PE antibodies. Total unselected MCF-7 cells can be divided into two distinct subpopulations: CD44+/CD24-, CD44-/CD24-. (B) CD44+/CD24- cells subpopulation of mammosphere culture cells. CD44+/CD24- cells and CD44-/CD24- cells were sorted by adding FITC anti-human CD24 and PE anti-human CD44 from mammosphere culture MCF-7 cells.
FITC=fluorescein isothiocyanate.
|
Cell cycle analysis
Experiments were performed with cells seeded at a density of 5×105 cells/dish (100 mmφ×20 mm high) in DMEM-F12 for 72 hours. Cell cycle analysis was performed using a bromo-deoxyuridine (BrdU) Flow Kit (Miltenyi Biotech Inc., Auburn, USA) with cells seeded in one of the dishes, after they were stimulated for 24 hours with 0, 10, 25, or 50 ng/mL bFGF in DMEM-F12. BrdU (10 µM) was added to the culture medium during each incubation period, and the cells were incubated for an additional 30 minutes. The cell cycle analysis was performed using a BrdU Flow Kit and a FACS Aria II flow cytometer (BD Biosciences). Data were analyzed using CellQuest software (BD Biosciences), and the percentages of cells in the G0/G1, S, and G2/M phases were determined (Figure 3).
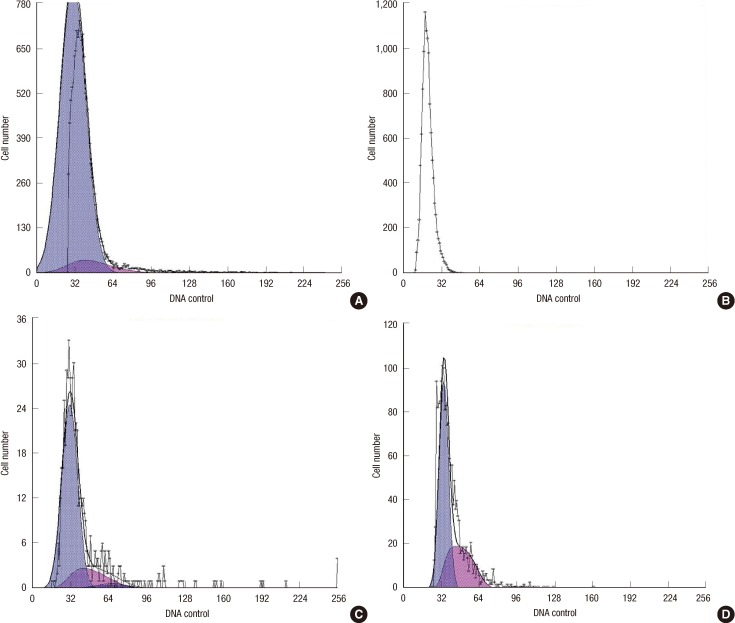 | Figure 3Cell cycle of CD44+/CD24- cells in different bFGF concentration. (A) Cell cycle distribution was measured by flow cytometry using a BrdU Flow Kit and the percent of the cell cycle phase is shown in a bar graph form with the G0/G1 (93.1%) and S (6.93%) phases for the cell lines in the absence of bFGF. (B) Cell cycle distribution was measured by flow cytometry using a BrdU Flow Kit and the percent of the cell cycle phase is shown in a bar graph form with the G0/G1 (85.3%) and S (14.7%) phases for the cell lines in the presence of bFGF (10 ng/mL). (C) Cell cycle distribution was measured by flow cytometry using a BrdU Flow Kit and the percent of the cell cycle phase is shown in a bar graph form with the G0/G1 (76.8%), G2 (4.34%) and S (18.8%) phases for the cell lines in the presence of bFGF (25 ng/mL). (D) Cell cycle distribution was measured by flow cytometry using a BrdU Flow Kit and the percent of the cell cycle phase is shown in a bar graph form with the G0/G1 (62.8%), and S (37.2%) phases for the cell lines in the presence of bFGF (50 ng/mL).
|
Go to :
RESULTS
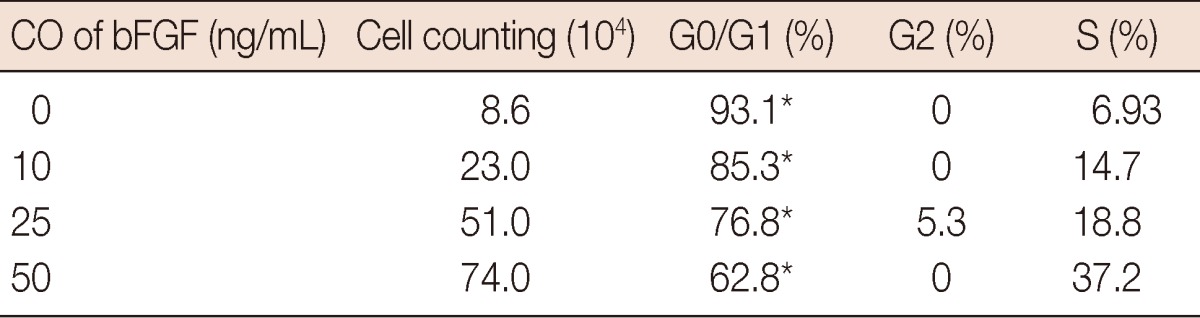
The percentage of cells in the G0/G1, S, and G2/M phases differed in the presence of bFGF. A cell cycle analysis was performed for 24 hours, then the cells were stimulated with DMEM-0.5, supplemented with bFGF (10, 25, or 50 ng/mL) to investigate the cell proliferation-specific effects of bFGF. As shown in Figure 1, the percentage of CD44+/CD24- cells in the G0/G1, S, and G2/M phases was significantly different under the different bFGF concentrations. The effect of bFGF on the cell cycle was quantified by bivariate BrdU flow cytometry analysis. Detailed values of the cell cycle analysis are summarized in Table 1. The percentage of cells in G0/G1 deceased with a change in bFGF concentration (Figure 4).
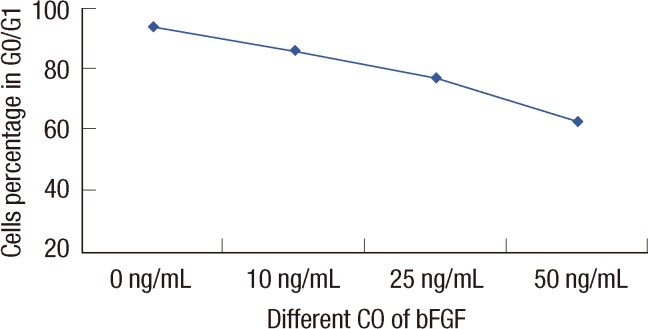 | Figure 4The correlation between the percentage of cells in G0/G1 under different bFGF concentration. The values were calculated on the correlation between the percentage of cells in G0/G1 under different bFGF concentration. Statistical differences were Z=2.28, p=0.023 by Cochran-Armitage. The percentage of cells in G0/G1 decreased with the bFGF concentration increased.
|
Table 1
The percentage of cells in G0/G1, S, and G2/M phases under different concertration of bFGF
Cell cycle analysis was performed at 24 hours after cells was stimulated with 0, 10, 25, 50 ng/mL basic fibroblast growth factor (bFGF) and cell numbers were determined. The percentage of cells in G0/G1 was greatly significant under different concentration of bFGF. The values were calculated from the cell numbers between 0, 10, 25, 50 ng/mL bFGF group. Statistical differences were χ2=18.08, *p=0.001.
![]()
Go to :
DISCUSSION
We demonstrated that the effect of bFGF on promoting CD44+/CD24-/low cell proliferation and progression through the G1/S phase transition was correlated with bFGF concentration.
FGF was first described in 1975 [8]. Only acidic and basic FGF were identified, based on the structural properties of FGF. Today, FGF proteins are understood to comprise a growth factor family that includes more than 20 proteins [9]. bFGF is mutagenic in various cell types, including bovine aortic endothelial cells [10] and adult rat schwann cells [11]; bFGF produces a 4-fold increase in DNA synthesis and a 3-fold rise in BrdU labeling, suggesting that it promotes the G1/S transition [12]. When added to culture medium, the proportion of nifedipine-reactive patient cells that underwent progression to the S and G2/M phases from the G0/G1 phase increased significantly compared to the proportion in nifedipine-non-reactive patient cells [13]; bFGF plays a concentration-dependent role regulating the cell cycle [12,14,15]. The effect of bFGF in the regulation of the MCF-7 cell cycle with CD44+/CD24- was evaluated in this study, as bFGF is a potential factor promoting the G1/S transition.
Breast cancer cells that express CD44 (CD44+) and show an absence or low levels of CD24-/low possess self-renewal capacity and the ability to initiate tumors in non-obese diabetic/severe combined immunodeficient mice from as few as 100 cells [16]. A recent study by Sheridan et al. [16] also showed that CD44+/CD24-/low breast cancer cells possess enhanced invasive properties. Cancer cells with CD44+/CD24- differentiate into two types of cell during malignancy development one type retains self-renewal capacity, often arresting in G0/G1, whereas the other type loses self-renewal capacity and differentiates into a cancer cell. Stem cells are mainly quiescent in the G0 state [14], which allows them to be drug-resistant. Some studies have shown that breast cancer cells with the CD44+/CD24- are more resistant to chemotherapy and radiotherapy than that of non-CSCs [3,4]. If we could promote cells to progress through the G1/S phase transition, breast cancer cell with CD44+/CD24 might be more easily destroyed. As the number of breast cancer cell with CD44+/CD24 was very low, a mammosphere culture was used to obtain more cancer-initiating cells by adding 5 µg/mL bovine insulin, 20 ng/mL bFGF-2, 10 ng/mL epidermal growth factor and 2% B-27 into the DMEM-F12. Then, CD44+/CD24- cells were sorted by flow cytometry, and 0.9% breast cancer cells with CD44+/CD24 were obtained from mammosphere culture, compared to 0.3% breast cancer cells with CD44+/CD24 obtained from normal MCF-7. Grimshaw et al. [17] found that mammosphere culture of pleural effusions enriched with CD44+/CD24low/- cells are capable of inducing tumors in severe combined immunodeficiency diseased mice.
Cell cycle analyses were performed to calculate the percentage of CD44+/CD24- cells in G0/G1, under different bFGF concentrations. The results showed that the percentage of cells in G0/G1 decreased as bFGF concentration increased (p=0.023). The proportion was higher in the presence of a high level of bFGF than that with a in the relatively low level of bFGF. When bFGF was added to the medium, cell proliferation advanced in CD44+/CD24-/low cells as well as in MCF-7, cells without being selected. Cell count increased with increasing bFGF concentration.
In summary, the findings indicate that: First, mammosphere cultures were able to enrich CD44+/CD24- cells; Second, bFGF was able to sustain CD44+/CD24- cell proliferation and promote cell progression through the G1/S phase transition; Third, this effect was correlated with bFGF concentration.
However, the mechanism for the role of bFGF to enable breast cancer cells with the CD44+/CD24- to undergo progression to the S and G2/M phases from the G0/G1 phase is unclear, thus, further studies must be conducted. Promoting breast cancer cell with the CD44+/CD24- marker through the G0/G1→G2/S phase transition may be an efficient way to destroy breast cancer cells.
Go to :
 XML Download
XML Download