

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The most frequently encountered malignancy of breast cancer tissue in women represents a classical model of estrogen dependent condition [1]. However, the effects of estrogen, like the other steroid hormones, are reported to be mediated due to their ability to cross the lipid bilayer membrane of cells and to bind the nuclear receptors [2], which in turn are reported to interact with a specific DNA sequence known as hormone responsive elements (HRE). The binding of the receptors to the HRE is reported to lead to the expression of the hormonal effects [2]. On the basis of presence of the nuclear estrogen receptors (ER), patients with breast cancer are classified into two groups: ER+ and ER-.
As in the case of estrogen, which is known to play a significant role in the development of human breast cancer, an anti-breast cancer protein called maspin (mammary serine proteinase inhibitor, 42 kDa) which is abundantly expressed in the normal mammary epithelial cells is reported toinhibit malignant breast cell invasion, promote apoptosis, and inhibit angiogenesis, and metastasis [3-5]. Studies have been conducted to determine the correlation between ER function and maspin synthesis if any, in the systemic development of breast cancer in female subjects, but the answer remains obscure.
We report herein the impairment of ER function in the synthesis of maspin through the impairment of estrogen-induced stimulation of nitric oxide (NO) synthesis in circulating neutrophils in the presence of human breast cancer in which the ER status of the malignant lesion was identical to that present in the patients' neutrophils.
METHODS
Ethical clearance
The protocol used in the study was approved by the Internal Review Board (IRB) of Sinha Institute of Medical Science and Technology. All of the patients with breast cancer and the age-matched normal female volunteers provided informed consent. Appropriate permission was also obtained from the IRB (approval no. 1/8/R/Br) for the use of rabbits in the studies.
Chemicals
Recombinant human maspin (rh maspin) was a kind gift of Dr. Sally Twining (Department of Biochemistry, Medical College of Wisconsin, Milwaukee, USA). The enzyme linked immunosorbent assay (ELISA) maxisorp plates were obtained from NUNC (Roskilde, Denmark). The estrogen and all other chemicals used were from Sigma Chemical Co. (St. Louis, USA). ERα and ERβ antibodies were obtained from Thermo Fisher Scientific (Rochester, USA).
Preparation of estrogen solution
Estrogen solution was prepared by dissolving the compound in 0.9% NaCl, and the pH was then adjusted to 7.4. The steroid hormone solution was used immediately, after preparation and discarded after use.
Selection of patients with breast cancer
Only female breast cancer patients or normal age-matched female volunteers 35 to 65 years of age (mean, 45 years; n=50), participated in the study. The occurrence of breast cancer was diagnosed by mammography or biopsy. None of the patients had received any therapy including radiation, surgery, or chemotherapy but were waiting for surgical intervention. These patients at presentation were staged as follows: Fifteen patients were staged at IIA (T1N1 M0), 20 patients were staged at IIB (T2N1 M0), 10 patients were staged at IIIA (T3N2 M0), and five patients were staged at IIIB (T4N0 M0). All subjects were M0. None of the subjects had a history of diabetes mellitus, systemic hypertension, severe infections, or life threatening cardiovascular or cerebrovascular conditions.
Selection of normal volunteers
An equal number of age-matched normal female volunteers in similar phases of their menstrual cycles as those of the selected breast cancer subjects were asked to participate in the study. These volunteers had never taken any contraceptives. All volunteers were asked to stop taking any medication including aspirin for at least 2 weeks before participating in the study.
Collection of blood
Blood samples (20-25 mL) were collected by venipuncture using siliconized 19-gauge needles in plastic vials, and anticoagulated by gentle mixing of 1 volume of 0.13 M sodium citrate with nine volumes of blood [6].
Immunization of the animals
Polyclonal antibodies against rh maspin were raised by repeated immunization of White New Zealand rabbits as described previously [7].
Preparation of neutrophil suspension and incubation of isolated neutrophils with estrogen
Neutrophils were isolated from the citrated blood samples as described previously [9]. The cell counts were determined using optical microscopy. The isolated neutrophils suspended in Hank's balanced salt solution (HBSS), pH 7.4, (6×109 cells/L) were incubated with different concentrations of estrogen as indicated for 4 hours at 37℃ under sterile conditions, and when needed, the nucleic acids were isolated from these incubated samples for in vitro translation of maspin as described below.
In vitro translation of maspin mRNA
Nucleic acids containing maspin mRNAs were isolated using the TRIzol method in neutrophils isolated from the blood samples [10]. The nucleic acid preparation was incubated with ribosomal preparation, a mixture of all amino acids (0.1 µmol each/mL) and 2 mM adenosine triphosphate as described previously [11]. After 6 hours of incubation under sterile conditions, the reaction mixture at 0℃ was centrifuged at 10,000×g for 10 minutes. The supernatant was used for the determination of maspin by ELISA as described below.
Preparation of ER immunohistochemistry slides of neutrophils for the determination of ER+ and ER- status
Isolated neutrophils were placed on glass slides, and frozen using cold liquid nitrogen vapor, and broken by the sliding of another glass slide over them to expose the nuclear receptors in the cells to the added fluorescent antibody. ER statuses were determined by immunohistochemical techniques using fluorescence tagged antibodies that recognized both α and β estrogen receptors [13]. The cells were then immediately observed and photographed under fluorescence microscopy.
Binding of estrogen to neutrophils
In preliminary experiments to determine the optimal time for estrogen binding, the normal neutrophil suspensions (6×109 cells/L) were incubated with 0.1 to 1.0 nM estrogen for different time periods at 37℃. The amounts of estrogen that bound to the neutrophils were determined after unbound hormone was separated from the bound hormone in the incubation mixture using ELISA as described below in the Scatchard plot analysis of estrogen binding.
Scatchard plot analysis of the equilibrium binding of estrogen to ER in neutrophils
The neutrophil suspensions were prepared from the blood samples from normal or from the subjects with breast cancer and suspended (6×109 cells/L) in HBSS buffer, pH 7.4, with different amounts of pure estrogen and incubated for different periods of time at 37℃. After incubation, the neutrophils with the bound estrogen were separated from the unbound hormone by filtration over a glass microfiber filter (GF/C; Sigma Chemical Co., St. Louis, USA) using a Millipore filter as described previously [14]. After filtration, the neutrophils were washed twice with equal volumes of HBSS buffer. The GF/C filter that retained the neutrophils with the bound hormone was subsequently air dried and estrogen was eluted from the filter by trituration with 1 mL of a CHCl3 CH3OH (1:1) mixture. After centrifugation at 0℃ and 5,000×g, portions of the supernatant were air dried. The air-dried sample was redissolved in 0.9% NaCl and its estrogen concentration was determined using ELISA. The results obtained were further verified using 1.0 µci (4-14C) estradiol (Tjaden Biosciences, Burlington, USA) to the incubation mixture. The bound estrogen was separated from the unbound ligand as described above and the radioactivity was measured to determine the binding using a scintillation counter as described previously [14].
Specific estrogen binding was determined by the addition of 10 mM unlabeled estrogen to the radio labeled estradiol as described above after subtracting the nonspecific binding from the total binding. The dissociation constant (Kd) and the receptor numbers (n) from the Scatchard plots [15] were determined by computer analysis.
Binding rate of (4-14C) estradiol to normal neutrophils in the presence of ERα and ERβ antibodies
In some study phases, the normal neutrophils suspension was incubated separately with ERα antibody or ERβ antibody or both in HBSS buffer as described above for 1 hour at 37℃ and the binding of estrogen was subsequently determined.
The rate of binding of (4-14C) estradiol to the normal neutrophils was determined after 1 hour of incubation (i.e., at the steady state of the binding). The bound 14C estradiol to the neutrophils was separated from the unbound compound via GF/C membrane filtration as described above. The radioactivity of the bound estrogen was then determined as described under Scatchard plots.
Statistical analyses
The obtained results are presented as mean±SD, while the significance of the results was determined using Student's t-test. Values of p<0.005 were considered significant. Where appropriate, the coefficient of correlation (r) of the results was also determined using the, Pearson test. GraphPad Prism software (GraphPad Software, San Diego, USA) was used for the statistical analyses.
RESULTS
Determination of ERα and ERβ breast cancer subtypes in normal, ER+, and ER- in peripheral neutrophils in blood
Immunohistochemical studies of the ER+ status in breast tissue and in the normal peripheral neutrophils demonstrated the presence of both ERα and ERβ subtypes. In contrast, ER- neutrophils from patients with breast cancer lacked the ERα and ERβ subtypes, suggesting similar pathophysiological defects in the synthesis of ER (α and ER β) proteins in both malignant breast tissues and peripheral neutrophils that were shown by histopathology to be identical to the malignant breast lesion subtype of the victim (Figure 1).
Equilibrium binding of estrogen to normal, ER+, and ER- neutrophils
Incubation of normal, ER+, and ER- neutrophils with 0.6 nM estrogen (the optimal estrogen concentration for maspin synthesis as determined in earlier experiments) produced saturable and specific binding profiles of the hormone to the neutrophils after 4 hours incubation at 37℃ at equilibrium (Figure 2). The amounts of estrogen bound to the ER+ neutrophils were, however, found to be markedly lower than those in the normal cells. In contrast, similar treatment of ER- neutrophils with estrogen showed little or no specific hormone binding, a finding that was similar to the nonspecific binding of the hormone to normal neutrophils (Figure 2).
Scatchard plot analysis of equilibrium binding of estrogen to neutrophils
A Scatchard plot of the equilibrium binding of estrogen to normal neutrophils produced a typical linear profile representing a homogeneous ER population within the neutrophils (Figure 3). Analysis of the equilibrium binding characteristics of estrogen demonstrated that the ER number in normal neutrophils was 4.18±1.02×107 sites/cell (Kd)=of 0.926 nM. A Scatchard plot of the equilibrium binding of estrogen to ER+ neutrophils also demonstrated a linear profile (Figure 3) similar to the normal neutrophil result. However, analysis of the hormone binding characteristics to ER+ neutrophils in the Scatchard plot showed that the ER count was nearly half (2.46±0.025×107 estrogen binding sites/cell) of that in normal neutrophils described above (p<0.001, n=5). However, the dissociation constants were identical between ER+ and normal neutrophils (Kd=0.926 nM). Since ER- neutrophils did not demonstrate estrogen binding, it was not possible to determine the ER number or the dissociation constant under the experimental conditions described in the Methods section.
Effect of different amounts of estrogen on nitric oxide synthesis in normal, ER+, and ER- neutrophils
Incubation of the normal neutrophils suspension in HBSS buffer, pH 7.4, for 4 hours with different amounts of estrogen as indicated at 37℃ resulted in NO synthesis in the incubation mixture (Figure 4). Both normal and ER+ neutrophils from the patients with breast cancer synthesized the most detectable NO at 0.6 nM estrogen. However, similar treatment of ER- neutrophils with different amounts of estrogen failed to produce any detectable amounts of NO in the reaction mixture under otherwise identical conditions.
We also found that normal neutrophils, which contained higher numbers of estrogen binding sites (4.18±1.02×107 sites/cell) than did ER+ neutrophils (2.46±0.025×107 sites/cell), synthesized higher quantities of NO (1.829±0.072 µM) than those synthesized by ER+ neutrophils (0.887±0.003 µM) when treated with 0.6 nM estrogen (p<0.001, n=15) (Figure 4).
These results indicated that the occurrence of estrogen-induced NO synthesis at the optimal concentration of the (i.e., 0.6 nM) was related to the number of ER both in normal and ER+ neutrophils, which had identical hormone affinities (Kd=0.926 nM) in both instances. In contrast, ER- neutrophils, which showed no estrogen binding, did not produce any NO upon estrogen treatment (Figure 4).
Effect of incubation of normal neutrophils with ERα or ERβ antibody on estrogen binding
The estrogen binding rate to normal neutrophils remained steady for 1 hour at 37℃ (19±0.12 nM/109 cells) but decreased to 17±0.11 nM/109 cells (11% inhibition) and 16±0.11 nM/109 cells (16% inhibition) upon incubation with ERβ and ERα antibody, respectively, as described in the Methods section. In contrast, when the normal neutrophils were incubated with both ERα and ERβ antibodies, estrogen binding decreased markedly (75%) to 0.5±0.11 nM/109 cells under otherwise identical conditions (Table 1).
Effect of estrogen-induced nitric oxide synthesis on maspin production in normal, ER+, and ER- neutrophils
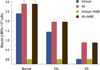
Since increased cellular concentrations of NO have been reported to result in increased maspin synthesis in neutrophils [6], experiments were conducted to determine the effect of estrogen-induced increase of NO in neutrophils in the synthesis of maspin in normal, ER+, and ER- neutrophils (Figure 5). We found that normal neutrophils in which NO synthesis was maximally stimulated at 0.6 nM also synthesized maximal levels of maspin (2.383±0.014 nM). In contrast, in ER+ neutrophils, in which the estrogen-induced synthesis of NO was nearly half, of that in normal cells, the amount of maspin synthesis was reduced to 1.422±0.029 nM (p<0.005) (Figures 4 and 5).
In contrast to normal and ER+ neutrophils, treatment of ER- neutrophils with similar amounts of estrogen failed to produce NO, or maspin (Figure 5). The addition of L-NG Nitroarginine Methyl Ester (NAME), an NO synthase inhibitor [16], to normal or ER+ neutrophils resulted in complete inhibition of the estrogen-induced synthesis of both NO and maspin.
Furthermore, treatment of normal and ER+ neutrophils with 5 µM NO (final) solution in 0.9% NaCl, instead of estrogen, produced 2.90±0.17 nM and 1.95±0.13 nM maspin, respectively. Interestingly, although the treatment of ER- neutrophils with estrogen failed to produce any NO or maspin, the addition of NO solution to the ER- cells resulted in the synthesis of 0.87±0.005 nM maspin even in the absence of added estrogen to the incubation mixture (p<0.005). The addition of NAME to the reaction mixture containing NO for the synthesis of maspin had no effect on the synthesis of the anti-breast cancer protein. These results indicated that although NO was capable of inducing maspin synthesis in ER- neutrophils, the addition of NAME, an inhibitor of nitric oxide synthase, had no effect on NO or the synthesis of maspin in the presence of added NO.
In other words, NAME had no effect on maspin synthesis induced by added NO to the reaction mixture containing neutrophils (Figure 6).
Correlation between estrogen-induced NO and maspin syntheses in normal and ER+ neutrophils
The coefficient of correlation "r" between the estrogen-induced NO synthesis and maspin production was +0.98 and +0.90 in normal and ER+ neutrophils, respectively, indicating that NO and maspin synthesis in these cells were highly and positively correlated.
DISCUSSION
The results of the current study demonstrated that ER status of the neutrophils in the peripheral blood of the subjects with breast cancer, as determined by immunohistochemical cytology, was identical to those of the patients' breast lesions of the victims (Figure 1).The ER activity was also demonstrated to be impaired in that steroid induced maspin synthesis due to impaired NO production occurred in both ER+ and ER- human breast cancer neutrophils compared to normal neutrophils (Figures 4 and 5).
The defective hormone receptor synthesis in cells other than in breast cancer cells themselves is not, unique, in fact, impaired insulin receptor activity has also been reported to occur both in breast cancer cells and in erythrocytes in the circulation in this condition [6]. It should be mentioned here that insulin, like estrogen, was also a potent inducer of maspin synthesis through NO production [6].
Furthermore, these results indicated that the estrogen-induced synthesis of maspin in neutrophils either from normal female volunteers or from patients with ER+ breast cancer was an NO dependent post ER interaction phenomenon. Although the treatment of both normal or ER+ neutrophils with estrogen resulted in the stimulation of maspin synthesis through NO production, similar treatment of ER- neutrophils with the hormone failed to stimulate either NO or maspin synthesis under identical conditions.
It is generally believed that steroid hormones including estrogen, which are lipid soluble, can freely pass through cell membrane bilayers of the cells [17] and are and subsequently bound to the nuclear receptors in the cytosol that mediate the expression of specific genes in the cells [6]. However, if estrogen were able to freely diffuse through the neutrophil membrane due to its lipid solubility of the steroid alone, then the synthesis of either NO or maspin might occur equally in ER+ and ER- neutrophils as determined by the effect of estrogen in the intact neutrophil suspensions (Figures 4 and 5). However, this was not the case. In contrast, our results suggested that the transport of estrogen into the neutrophils was an ER dependent physiological event. In other words, the ERs played an essential role in the transport of the steroid hormone into the cell through the lipid membrane bilayers and the transportation process of estrogen was not necessarily due only to the lipid soluble nature of the steroid hormone. This inference was made due to immunohistological studies that indicated the presence of both ERα and ERβ in the normal and ER+ neutrophils and the absence of both receptor subtypes in ER- neutrophils (Figure 1). These results suggested that both ERα and ERβ could be involved in the transport of estrogen into neutrophils.
The above conclusion was also supported by the fact that the incubation of normal neutrophils with ERα, ERβ, or both antibodies inhibited estrogen binding. We found that although incubation of the normal neutrophils suspension with ERα antibody only blocked estrogen binding by 15%, incubation of these cells with ERβ antibody blocked the hormone binding by 11%. In contrast, incubation of the normal neutrophils with both ERα and ERβ together blocked the binding by 75%.
Although the above results suggested that both ERα and ERβ could play an essential role in the binding/transportation of estrogen in neutrophils, the role of ERα and ERβ in estrogen-induced maspin synthesis via NO production could be of lesser importance, at least in the case of estrogen-induced synthesis of maspin in neutrophils (Figure 5). It was found that while the treatment of normal neutrophils with 25 nM progesterone instead of 0.6 nM estrogen increased the NO synthesis from 0.544 to 1.317 µM with simultaneous increase of maspin from 1.36 to 2.33 nM maspin/109 cells, similar treatment of ER+ neutrophils with 25 nM progesterone increased the NO synthesis from 0.32 to 0.73 µM and increased maspin synthesis from 0.41 to 1.14 nM/109 cells (data not shown). It could, however, be argued that progesterone at higher concentrations was simply mimicking estrogen in vitro. The physiological level of progesterone is also known to be 80 fold higher than that of estrogen in women in the mid cycle of their menstruation periods.
Although the effects of progesterone and estrogen could represent similar effects of two different hormones in the synthesis of maspin via NO production, these results nevertheless might also suggest favorably comparable effects of these two steroid hormones in the control of the development of human breast cancer through NO induced maspin production in the system instead of antagonizing their effects in the control of the condition as has been reported previously [18].
It could be argued that the lack of maspin synthesis in ER- neutrophils was not due to the lack of estrogen receptors in the cells (Figures 4 and 5) but rather was due to mutation of the gene involved in the synthesis of the anti breast cancer protein in these neutrophils obtained from patients with breast cancer. However, the direct addition of NO solution to the ER- neutrophils that did synthesize maspin due to the presence of NO suggested that the estrogen-induced impairment of maspin synthesis in the ER- neutrophils might not be directly related to the mutation of the maspin gene because NO could not stimulate maspin synthesis in ER- neutrophils.
On the other hand, these results indicated that as a result of estrogen binding to its nuclear receptors, the synthesis of NO synthase occurred due to the nuclear receptor interaction with the HRE in the DNA of the neutrophils. NO formed due to the catalytic activity of the NO synthase produced by the neutrophils and, subsequently stimulated maspin synthesis. The ability of the formed NO to synthesize maspin in neutrophils has been reported before [6].
The inhibition of estrogen-induced NO synthase by NAME inhibited maspin synthesis. On the other hand, treatment of the neutrophils with NO solution resulted in maspin synthesis, of even in the presence of NAME or in the absence of estrogen. These results suggested that in the estrogen-induced maspin production due to the activation of ER that stimulated NO synthesis, the NO was acting like a "messenger" for maspin synthesis in neutrophils.
It has been reported previously that the prognostic outcome of breast cancer with ER- lesions was worse than that with ER+ lesions [19,20].
Our results, as presented above, suggested that estrogen-induced maspin synthesis was severely impaired in ER- neutrophils compared to that in ER+ neutrophils, as such, impaired the estrogen-induced maspin synthesis in patients with ER- breast cancer might result in worse prognostic outcome when the lesions are ER- status than when the lesions are ER+ status.

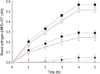
 ) represent specific binding, while the dotted lines (
) represent specific binding, while the dotted lines ( ) represent total binding of estrogen to the neutrophils. The results are the mean±SD of five different experiments each in triplicate using blood of five different patients with ER+ or ER- breast cancer or from normal volunteers.
) represent total binding of estrogen to the neutrophils. The results are the mean±SD of five different experiments each in triplicate using blood of five different patients with ER+ or ER- breast cancer or from normal volunteers.



 XML Download
XML Download