

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Clinically, breast cancer develops through sequential stages from normal ductal epithelium to hyperplasia, ductal carcinoma in situ (DCIS), invasive cancer, and metastatic carcinoma. Although extensive research has focused on identifying the molecular changes involved in carcinogenesis, the mechanisms underlying the development of breast cancer remain unclear [1]. Various reports have demonstrated that the expression of Annexin A1 (ANXA1) is ordinarily regulated in the mammary gland during the developmental period. Although the molecular action of ANXA1 is not yet fully understood, ANXA1 appears to take part in intracellular signaling and cell differentiation. ANXA1 is a 37-kDa calcium- and phospholipid-binding protein of the annexin superfamily, and has been detected in miscellaneous organisms, including vertebrates, invertebrates, and plants [2]. Further, ANXA1 is an important mediator in glucocorticoid-regulated inflammatory responses that modulates the activation of innate immune cells, such as neutrophils and macrophages [3], and it is known to have an association with dexamethasone-induced cell growth arrest [4]. The ANXA1 protein is partially located in the nucleus of endothelial cells, which indicates that it has a role in an intracellular signaling pathway. Because ANXA1 is distinctively expressed in the mammary gland during embryonic development [5-7], we postulate an association between ANXA1 and breast cancer development. Decreased expression of ANXA1 has been consistently reported at both the RNA and protein levels in breast cancer [8,9]; however, the role of ANXA1 expression in tumor initiation or progression has remained unclear. In this study, we specified the patterns of ANXA1 expression in various breast diseases including benign tumors and malignancies and then analyzed the relationship with prognostic factors and survivals of ANXA1 in invasive breast cancer.
METHODS
Patients
Between April 2005 and December 2007, human breast tissue samples from benign or malignant lesions were retrieved from patients who had undergone breast surgery or core-needle biopsy at the Ewha Medical Center. Normal breast tissues were obtained from non-tumorous areas of breast cancer patients more than 10 cm from the primary cancer. All of the patients who were enrolled in this study gave written informed consent in accordance with the Declaration of Helsinki. The study was reviewed and approved by the local institutional review committees.
Immunohistochemistry
The patterns of ANXA1 expression were examined by immunohistochemical (IHC) staining. Breast tissues were formalin-fixed and paraffin-embedded using standard methods. Monoclonal anti-ANXA1 antibody (catalog no. 610067; BD Transduction Laboratories, Lexington, USA), a specific antibody for ANXA1, has been suitable for immunochemical use on a variety of formalin-fixed and paraffin-embedded tissues of bovine and human origin [10,11]. Monoclonal anti-ANXA antibody was added as a 100 mL aliquot at a concentration of 1 mg/mL. For isotype control monoclonal antibody (mAb), we used a purified mouse IgG1.kappa (MOPC 21) from Sigma Chemical (St. Louis, USA). Tissue sections were cut in 4 µm slices and mounted on protein-coated glass slides. After dewaxing in xylene and rehydration in a series of alcohols, the staining procedure was carried out as follows. All the linking and labeling reagents, substrates, and chromogens were supplied from BioGenex (San Ramon, USA) as a super-sensitive immunodetection system. All procedures were carried out at room temperature. Before the mouse mAb was applied, a sufficient amount of protein blocking reagent (normal goat serum), which completely covered each tissue section, was placed on the slides for 20 minutes. After blotting around each section, primary mAb (1:100 dilution) was applied to the slides, incubated for 60 minutes, followed by the addition of prediluted, biotinylated anti-immunoglobulin for 20 minutes, and a labeling agent (alkaline phosphatase-labeled streptavidin) for 20 minutes [12]. For the chromagen, one tablet of Fast Red substrate was dissolved in 5 mL of naphtol phosphate-substrate solution, and levamisole was added to the substrate solution to block endogenous alkaline phosphatase activity. Enough the chromogen substrate solution was added to cover each section entirely, and the slides were incubated for 10-30 minutes, or until acceptable color intensity developed. Finally, the slides were counterstained with Mayer's hematoxylin solution and mounted with aqueous mounting medium (BioGenex). Each tissue section was analysed and scored by a pathologist. ANXA1 expression was scored as follows: the staining intensity was evaluated from 0 to 3 (representing negative to strong staining, respectively) and % cells in each intensity was obtained. The overall score was determined as follows: overall score=[(% cells with visual score 1)×1]+[(% cells with visual score 2)×2]+[(% cells with visual score 3)×3]; expression was positive if the score was more than 70. The hormone receptors (HR) (either., estrogen receptor or progesterone receptor) were considered positive when ≥10% of positive tumor cells had nuclear staining. Membranous staining for HER2 was scored as follows: 0, no staining or membranous staining in <10% of the cells; 1+, faint, incomplete staining in 10% of the cells; 2+, weak-to-moderate complete staining in 10% of the cells; and 3+, strong complete staining in 10% of the cells. HER2 overexpression was defined as a score of 3+. p53 was considered positive if >10% of the cells were stained with strong intensity.
Statistical analysis
Statistical analysis was performed with SPSS version 17.0 (SPSS Inc., Chicago, USA). Pearson's χ2 test was used to determine the correlation between variables, and the Cox proportional regression hazard model was employed with several variables. Kaplan-Meier curves were plotted from breast cancer-related survival (BCRS) and relapse-free survival (RFS) data. Univariate and multivariate p-values were two-sided and were considered to be significant when they were equal to or less than 0.05.
RESULTS
Cilinicopathologic characteristics of patients and immunohistochemical results of ANXA1
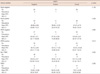
One hundred eighty-two cases were enrolled in this study, and the mean age of all female patients was 46.34±11.5 years. ANXA1 was strongly expressed in myoepithelial cells compared to epithelial cells in normal breast tissues (p<0.001) (Figure 1A). ANXA1 positivity was 87.7% in normal tissues, 53.2% in benign diseases, 14.3% in DCIS, and 30.9% in invasive carcinoma. A significant loss of ANXA1 expression was observed both in DCIS (n=7) and invasive carcinoma (n=75, Figure 1B) compared to normal epithelium (n=73, p<0.001) and benign breast diseases (n=27, p<0.001) including intraductal papilloma (n=10), sclerosinng adenosis (n=4), atypical ductal hyperplasia (n=2), fibroadenoma (n=6), and fibrocystic change (n=5). There was no difference between the ANXA1 expression of DCIS and invasive carcinoma (p=1.000). In the analysis within invasive breast carcinoma, there was no significant difference in ANXA1 expression according to age (p= 0.458), tumor size (p=1.000), nodal status (p=1.000), HER2 overexpression (p=0.586), p53 expression (p=0.387), histologic grade (p=0.347), or recurrence (p=0.084). ANXA1 expression was significantly correlated with unfavorable prognostic factors including HR negativity (p<0.001), HER2 type and triple negative breast cancer (TNBC) (p<0.001), and high nuclear grade (p=0.004) (Table 1). Althougth the data has not been shown in a table, there was no difference in ANXA1 expression among histologic subtypes of invasive breast cancer (p=0.520).
Relationship between ANXA1 expression and clinical outcomes
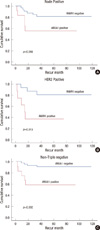
The median follow-up duration was 63.0±24.08 months (range, 6.0-117.0) for invasive ductal carcinoma (n=75). In a univariate analysis, HER2 overexpression (p=0.022) and advanced stage (p=0.031) were correlated with poor BCRS (Table 2). Although there was no statistical significance, ANXA1 expression tended to be associated with unfavorable BCRS (p= 0.062). In relapse-free survival (RFS), nodal status (p=0.047), HR status (p=0.004), HER2 overexpression (p=0.012), TNM stage (p=0.001), and nuclear grade (p=0.007) had the statistical significance. In contrast, there was no significant difference in BCRS and RFS according to ANXA1 expression in the multivariate analysis (Table 3). HER2 positivity and advanced stage were associated with poor RFS in multivariate analysis (p=0.037 and p=0.048, respectively).
DISCUSSION
A reduction in ANXA1 expression in breast cancer has been consistently reported [8,9], a finding which was corroborated by this study. We also identified that ANXA1 expression was more prominent in myoepithelium than in epithelium in normal breast tissue; however, disappearance of myoepithelial cells has been a major hallmark in invasive breast cancer [13]. Shen et al. [1] tried to determine whether the decreased integrated maximal intensity of ANXA1 staining observed in DCIS samples was simply due to a reduction in the number of these cells, or if there was an underlying mechanism involved, such as a decreased level or frequency of cellular ANXA1 protein expression. They examined the expression of ANXA1 in myoepithelial cells from the TMA spots of normal, hyperplastic, and DCIS, and any minor expression was included. They concluded that the proportion of ANXA1 expression decreased, and the proportion of ANXA1 non-expressing areas increased in myoepithelial cells of non-malignant tissues compared to DCIS lesions. However, the distribution of ANXA1 expression in myoepithelial cells did not reveal notable differences between normal and hyperplastic ductal lesions. These results indicated that the loss of ANXA1 expression occurs early during malignant transformation. In this study, we found a significant loss of ANXA1 expression both in DCIS and invasive carcinoma compared to normal epithelium and benign breast diseases which supports previous studies. There have been a number of theories about the potential mechanism of ANXA1 in cancer development. For example, ANXA1 has been linked with reduced cell proliferation involving the regulation of the extracellular signal-regulated kinase (ERK)/mitogen-activated protein kinase (MAPK) signal transduction pathway. Forced expression of ANXA1 activates the ERK/MAPK pathway and reduces cell proliferation by disrupting the actin skeleton and ablating cyclin D1 expression [14,15]. Multiple members of the annexin family, including annexins 1, 2, 5, 6, and 7, have been reported to have involvement in the down-regulation of cell proliferation. Therefore ANXA1 has been postulated to have tumor suppressing properties [2,16].
On the contrary, some studies have shown that a higher level of ANXA1 in lymph node metastasis in comparison with primary breast cancer [14]. A similar finding was observed by Pencil and Toth [17] in rat metastatic mammary cancer and by Ahn et al. [18] in human breast cancer. Further, ANXA1 expression was found to be down-regulated by breast cancer metastasis suppressor 1 gene in the MDA-MB-435 breast cancer cell line metastatic to the lung in an athymic mouse model [19]. A more recent study showed that membrane ANXA1 had an autocrine/paracrine action in stimulating the migration of SKCO-15 colorectal cancer cells through activation of n-formyl peptide receptors, which is a leukocyte migratory factor [20]. A recent study [21] showed that ANXA1 expression provoked drug resistance in breast cancer cells. The overexpression of ANXA1 induced resistance to anti-cancer drugs, and ANXA1-depleted tumor cells showed increased sensitivity to anticancer drugs. Therefore, these results supported that ANXA1 also has oncogenic potential for breast cancer progression and metastasis via multiple mechanisms. In the current study, ANXA1 expression was significantly correlated with unfavorable prognostic features of breast cancer, such as hormone receptor negativity (p<0.001), positive HER2 status and TNBC type (p<0.001), high nuclear grade (p=0.004). The ANXA1 positivity was also correlated with poor RFS in some groups of invasive breast cancer such as node positive, HER2 positive and non-TNBC. This results could be explained that, in condition with unfavorable factors including node positive and HER2 positive status, ANXA1 might tend to re-express in cancer tissue and which result in more metastatic and aggressive phenotype. Given that this study had a limited number of patients, further study may be required to further support out findings.
Although large scale, solid base studies are required to draw clinically meaningful conclusions, we found a major loss of ANXA1 in breast cancer tissues, including DCIS and invasive carcinoma, relative to normal and benign breast tissues. However, in cases of invasive breast cancer, ANXA1 positivity was significantly correlated with unfavorable prognostic factors. These results implied that ANXA1 plays dualistic roles and mechanisms in cancer development and progression in terms of both tumor suppressive and oncogenic activity.




 XML Download
XML Download