

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Pomegranate (Punica granatum) is an ancient fruit that has been used as a medicine in many cultural areas, and nowadays it is widely consumed as fresh fruits or processed into such forms as juice, jam, and wine in west and asian countries.(1) It has anti-inflammatory effects, anti-oxidant effects, anti-angiogenic potential and anti-proliferative effects.(2-4) Epidemiologically, some reports showed that the extract of pomegranate has chemopreventive and anti-cancer effect on breast cancer.(5)
Pomegranate contains numerous phytochemicals such as ellagic acid, quercetin, kaempferol and luteolin glycosides.(6,7) Among these compounds, ellagic aicd has been demonstrated to have antitumor and anti-apoptotic effect in bladder cancer cells, prostate cancer cells, cervical cancer cells, colon cancer cells, oral cancer cells and leukemic cells.(4,8-10) Only a few investigators reported about the relation between ellagic acid and breast cancer. However the effect of ellagic acid on breast cancer is still not well understood.(6,11-12)
The mechanism by ellagic acid which elicits anti-proliferative effects still remains poorly understood, although it has been found that the apoptosis caused by ellagic acid is associated with p53/p21 expression, cell cycle arrest, regulating matrix metalloproteinases, vascular endothelial growth factor expression and platelet-derived growth factor.(9,13,14) Other studies demonstrated that ellagic acid provoked the induction of apoptosis via intrinsic pathway through bcl-xL down-regulation with mitochondrial release of cytochrome c into the cytosol.(9,15)
Our objective in this study was to evaluate the biological activity of ellagic acid by comparing the anti-proliferative effect and apoptotic pathway of ellagic acid between ER positive MCF-7 and ER negative MDA-MB-231 human breast cancer cell lines.
METHODS
Chemicals and reagents
Ellagic acid was supplied from Calbiochem (San Diego, USA). Fetal bovine serum (FBS) was obtained from JRH Bioscience, Inc. (St. Lenexa, USA). Other chemicals, unless otherwise specified, were purchased from Sigma Chemical Co. (St. Louis, USA).
Breast cancer cell lines and culture method
ER-positive MCF-7 and ER-negative MDA-MB-231 human breast cancer cells lines were obtained from the Korean Cell Link Bank (Seoul, Korea).
These were cultured in Dulbecco's modified Eagle's medium (DMEM) (Gibco BRL, Thessalonica, Greece) supplemented with 10% fetal bovine serum in an incubator with 5% CO2 under 37℃. For experiments, cells were cultured for 3 days in phenol red free DMEM (Gibco BRL, Thessalonica, Greece) and 5% (v/v) charcoal-dextran stripped FBS (CSS; Hyclone, Logon, USA) prior to use and they were maintained in the same media. Cell suspensions were diluted to a cell density of 5×105 cells/cm2 into a 96-well plate for 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. For western blotting, cancer cells were cultured in 100-mm tissue culture Petri dishes. The following day, chemicals dissolved in DMSO were added. The final DMSO concentration did not exceed 0.3% (v/v). They were incubated in a 5% CO2 atmosphere and 100% humidity.
Cell proliferation assay
The MTT colorimetric assay was performed to identify tumor cell growth after treatment of each concentration (0, 10, 25, 50, 100, 250 µM) of ellagic acid. Plates were incubated in the presence of MTT dye for 4 hr. Mitochondrial dehydrogenase activity reduced the yellow MTT dye to a purple formazan, which was then solubilized with acidified isopropanol and absorbance was read at 570 nm on an enzyme-linked immunosorbent assay (ELISA) plate reader (Molecular Devices Inc., Union city, USA).
Cell count
Cell growth was also assessed by cell count. Cells were plated in 100-mm tissue culture Petri dishes and were cultured by the method as mentioned above. After 24 hr of incubation, cells were dislodged from the vessel bottom by treatment with a trypsin-EDTA solution. After vigorous pipetting, the cell numbers were measured by counting under a light microscope using a Neubauer hemocytometer.
Cell cycle analysis
Cell cycle phase distribution was analyzed by flow cytometry (Becton-Dickinson Immunocytometry System, Franklin Lakes, USA). Cultures were incubated with each dose of ellagic acid for 24 hr and harvested. The cells were fixed in 70% ethanol at -20℃ overnight. Cells were washed twice with PBS and stained with PBS containing 40 µg/mL propidium iodide, 100 µg/mL RNase (Sigma, St. Louis, USA) and 0.1% triton x-100. After 30 min at 37℃ in the dark, the cells were analyzed with flow cytometry equipped with an argon laser at 488 nm. Distribution of cells in different phases of cell cycle was determined using a software program Modfit (Becton-Dickinson Immunocytometry System).
Western blot analysis for bcl-xL and survivin
To harvest the cells, they were washed twice with PBS and scraped from culture dishes with lysis buffer containing 50 mM Tris-Cl (pH7.4), 150 mM NaCl, 1% Triton X-100, 0.1 mM DTT, 1mM EDTA, 1 mM PMSF and 1 µg/mL aprotinin. Cell lysates were centrifuged at 10,000×g for 10 min. After total protein extraction, 20 µg of proteins were separated by SDS-PAGE on 10% acrylamide Laemmli gels and transferred onto nitrocellulose membranes overnight. Membranes were blocked with the primary antibody for 90 min with 3% nonfat milk in PBS containing 0.5% Tween 20 and incubated for 90 min at room temperature with bcl-xL (R&D systems Inc., USA), survivin (Santa Cruz Biotechnology Inc., Santa Cruz, USA), pS2 (Santa Cruz Biotechnology Inc., Santa Cruz, USA) and c-fos (Calbiochem, San Diego, USA) antibodies. After incubation with peroxidase-conjugated secondary antibody for 90 min at room temperature, blots were developed using the ECL detection system (Amersham Biosciences, Uppsala, Sweden). The results of western blotting were standardized with the density of β-actin.
Cell fractionation and western blot analysis for cytochrome c
Mitochondria and cytosolic fractions were prepared using a Mitochondria fractionation kit (Active Motif, Carlsbad, USA) according to the manufacturer's recommendations and other investigators who used this kit.(16,17) Western blotting was performed using the above method with cytochrome c antibody (Santa Cruz Biotechnology Inc., Santa Cruz, USA).
RESULTS
Effect of ellagic acid on proliferation of MCF-7 and MDA-MB-231 cell lines
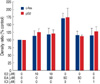
The concentration-dependent effects of ellagic acid on proliferation of MBA-MB-231 and MCF-7 human breast cancer cell lines are shown in Figure 1. In MDA-MB-231 cell lines, ellagic acid showed significant anti-proliferative effects with dose dependent manner. In MCF-7 cells, the anti-proliferative effect was observed in only high concentration (more than 250 µM).
Figure 2 represents that anti-proliferative effect of ellagic acid on MDA-MB-231 cells is dose-time dependent pattern. However, there was no time dependent anti-proliferation effect in MCF-7 cells.
Cell count showed the same result with MTT assay (data is not shown).
Effect of ellagic acid on cell cycle kinetics
The effects of ellagic acid on cell cycle in MDA-MB-231 cell line and MCF-7 cell line were determined using flow cytometry. The results for cell cycle analysis indicated that, during the 24 hr time period, ellagic acid has no influence on the change of cell cycle on both type of cell lines (Figure 3).
Effect of ellagic acid on expression of bcl-xL, cytochrome c and survivin
Ellagic acid decreased expression of bcl-xL and increased mitochondrial release of cytochrome c on MDA-MB-231 cells according to the increase of concentration. In addition, expression of survivin in MDA-MB-231 cells decreased with dose-dependent pattern.
In MCF-7 cells, expressions of bcl-xL and cytochrome c were not changed and expression of survivin was decreased with dose-dependent pattern. Although the expression of survivin in MCF-7 was decreased with stimulation of ellagic acid, the range of decrease was lesser than that in MDA-MB-231 (Figure 4).
It means that the anti-proliferative effect of ellagic acid appears dominantly through apoptotic pathway, especially decrease of bcl-xL and survivin, in MDA-MB-231 cells. Although decrease of expression of survivin was observed also in MCF-7, the stimulation of apoptotic pathway by ellagic acid was not enough to induce the anti-proliferative effect in MCF-7 cells.
Effect of ellagic acid on expression of c-fos and pS2
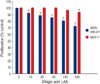
Expression of c-fos and pS2 in MCF-7 cells after 48 hr of exposure to E2, ellagic acid and ICI182,780 were evaluated to investigate whether ellagic acid could stimulate ER or not. Ellagic acid was able to induce an up-regulation of c-fos and pS2 protein (Figure 5). It induced pS2 protein with 1.39-fold and c-fos with 1.51-fold more than E2. The expression of pS2 and c-fos by ellagic acid was blocked by co-treatment of cells with ICI182,780.
DISCUSSION
Ellagic acid inhibited proliferation of MDA-MB-231 cells, but the effect on the proliferation of MCF-7 cells was shown only in high concentration. These anti-proliferative effects of ellagic acid in two types of human breast cancer cell lines were very distinguishable. Anti-proliferation effect of ellagic acid has been studied in many kind of cancer cell lines although the exact mechanism has not been well investigated.(4,18-21) Many investigators reported that ellagic acid represents anti-proliferative activity through the induction of cell cycle arrest, apoptosis or antioxidant effects.(6,9,22)
Cell cycle arrest that is induced by ellagic acid is not definite and still controversial. Some studies showed that ellagic acid induced G0/G1 phase arrest in cervical epithelial carcinoma cell, leukemic cells, bladder cancer cells and colorectal adenocarcinoma cells.(9,23) Another studies reported that it caused S phase arrest.(15) The other investigator informed that ellagic acid has no significant influence on cell cycle.(9) In present study, cell cycle was not changed by ellagic acid in both MDA-MB-231 cells and MCF-7 cells.
Although the influence of ellagic acid on cell cycle is not definite, most of reports agreed that ellagic acid induces apoptosis in various types of cancer cells.(15,23) In general, two major pathways are involved in apoptosis. One is the mitochondrion-initiated pathway and the other is the cell surface death receptor pathway. A report showed that ellagic acid induces apoptosis via mitochondrial-initiated pathway in colon cancer cells. It showed that this phenomenon occurs through bcl-xL down-regulation with mitochondrial release of cytochrome c into the cytosol activation of initiator caspase 9 and effecter caspase 3.(15) Bcl-xL belongs to the antiapoptotic proteins of the bcl-2 family, and decrease of bcl-xL stimulates release of cytochrome c from mitochondria. The released cytochrome c activates caspase 3 and it induces apoptosis. In our study, the expression of bcl-xL in MDA-MB-231 was definitely decreased with the increase of concentration of ellagic acid. In addition, cytochrome c in cytosol increased with the decrease the expression of bcl-xL. However, expressions of bcl-xL and cytochrome c in MCF-7 cells were not changed after treatment of ellagic acid even in high dose. It seems that the expression of bcl-xL might have the relation with the different response of both cell lines to ellagic acid. Other studies also reported that down regulation of bcl-xL protein expression in various tumor cell lines resulted in activation of apoptosis and decreased cellular proliferation.(24,25)
The expression of survivin, which belongs to IAP family was also examined. IAP family can bind to the cell death proteases such as caspase 3 and caspase 7 and inhibits caspase activity and cell death in cells exposed to apoptotic stimuli.(26,27) In breast cancer, survivin is a significant predictor of survival in patients with poor prognostic breast cancer.(28) Our result presents that ellagic acid decreases survivin in both human breast cancer cell lines. This indicates that one of targets of ellagic acid is IAP family, although the process is not exactly same in both cells. Decreased amount of survivin expression were larger in MDA-MB-231 cell than that in MCF-7.
When we focused on breast cancer, there were only few studies of the role of ellagic acid. Recently, anti-angiogenesis effect of ellagic acid on breast cancer cell line was reported.(29) However, most of the papers were only about the chemoprevention effect of ellagic acid on breast cancer. Almost all investigators experienced using various cancer cell lines-such as colorectal cancer, prostate cancer or lung cancer. MCF-7 was only added as one of these cancer cell lines. We hoped to avoid the bias that ellagic acid is less effective compared with other kind of cancer, and studied the antiproliferative effect of ellagic acid on MDA-MB-231 and MCF-7 which have been used most commonly as breast cancer cell lines. The results showed that if we could find specific marker for ellagic acid, it could show superior effect on some portion of breast cancers.
The limitation of this study is the small number of cancer cell lines that were experienced. Even though MDA-MB-231 cells and MCF-7 cells are most frequently studied cell lines, they can not represent all characteristics of breast cancer. Experiences using various breast cancer cell lines might find the specific target of ellagic acid, and this pioneer study could be helpful to investigate this field.
To investigate whether the different effect of ellagic acid in these breast cancer cell lines has any relation with estrogen receptor, we studied the expression of c-fos and pS2 in MCF-7 cells treated with ellagic acid. c-fos and pS2 has been examined for sensitivity to ligand structure in MCF-7 cells.(30) Ellagic acid more powerfully stimulated the expression of c-fos and pS2 rather than E2 and these expressions were inhibited by ER antagonist, ICI 182,780. Putting these various result together, ellagic acid might have an anti-estrogenic effect in MCF-7 cells. Interestingly, although ellagic acid inhibits the cell proliferation induced by E2, expression of c-fos and pS2 treated with ellagic acid is much more increased than E2 treated one. Therefore, the effect of ellagic acid could be appeared mainly through ER in MCF-7.
CONCLUSION
Our results indicate that ellagic acid has anti-proliferative effect in MDA-MB-231 cells. This effect of ellagic acid appears through apoptotic pathway such as the decrease of expression of bcl-xL and decrease of expression of survivin that are members of IAP family. Although there was no definitive anti-proliferative effect in MCF-7 cells, suppression of expression of survivin were observed also. These data suggest that ellagic acid has different effects on each breast cancer cell line. If specific target of ellagic acid could be identified in breast cancer, ellagic acid may be used as a chemotherapeutic agent for the treatment of breast cancer.



 XML Download
XML Download