

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) is an S. aureus that became resistant to β-lactam antibiotics by acquiring mecA gene on its chromosomal DNA [1]. MRSA was first isolated in 1960 in England [2], and became a worldwide epidemic since 1970s. At least three distinct genotypes of MRSA were present in the 1980s [1], and two of them are still prevalent in the world as multi-drug-resistant healthcare-associated MRSA (HA-MRSA) [3]. They acquired resistance to practically all the antibiotics introduced into clinical use in the past half century. They conquered even the 'last-resort' antibiotic vancomycin [4, 5]. On top of it, from the early 1990s, MRSA with non-multi-drug-resistance phenotype emerged from various countries of the world such as Australia [6, 7], the USA [8], and France [9]. These MRSA strains appeared to have emerged outside the hospital and designated community-associated (or acquired) MRSA (CA-MRSA). The year 2010 marked the 50th anniversary of our continuous struggle with MRSA. However, with an unexpected new surge of MRSA strains in both hospital and community, we are forced to fight an even bitterer fight with them. In this review, we describe the genetic feature of the organism to lay a basis for our future project to control MRSA.
Go to : 
S. aureus genome
1. Genomic islands (GIs)
S. aureus possesses several GIs on the chromosome. Figure 1A illustrates comparison of the genome structures of 14 S. aureus strains. All open reading frames (orfs) identified on their chromosomes are shown as vertical lines, in which 1887 (denoted by grey lines) are commonly shared by the 14 strains. Red lines indicate the orfs that are not present in all the strains. On each chromosome there are regions where grey lines are sparse and red lines predominate. Those regions correspond to GIs. The genes essential for the life of bacteria are all included in the grey orfs. In contrast, GIs include genes associated with virulence or useful genes for some specific environmental challenges (e.g. drug-resistance gene), which presumably were acquired from other organisms by horizontal gene transfer. These S. aureus GIs include integrated prophages, oriC environ (discussed later) and νSa islands (ν stands for island) [10]. The S. aureus GIs are mostly comprised of exogenous genes acquired by intra-species gene transfer [11]. Since S. aureus is by far the most virulent among various staphylococcal species, the GIs contain many virulence genes such as tsst-1 encoding toxic shock syndrome toxin, and multiple enterotoxin (se) genes. There are two classes in the νSa islands: the first class is designated νSan where n is an Arabic numeral indicating the position in the S. aureus chromosome where it is localized. They correspond to the integration sites for the mobile pathogenicity islands of Staphylococcus (SaPI) and a mobile resistant island (SaRI) [12-14]. The latter nomenclature is based on the unique genetic structure common to these mobile genetic elements. Correlation of the two nomenclature systems is summarized in Table 1. As shown in Figure 1A, νSan classification refers to the location of the attachment site of the islands, and is correlated well with the evolutionary tree for the SaPI integrase gene (Fig. 1B).
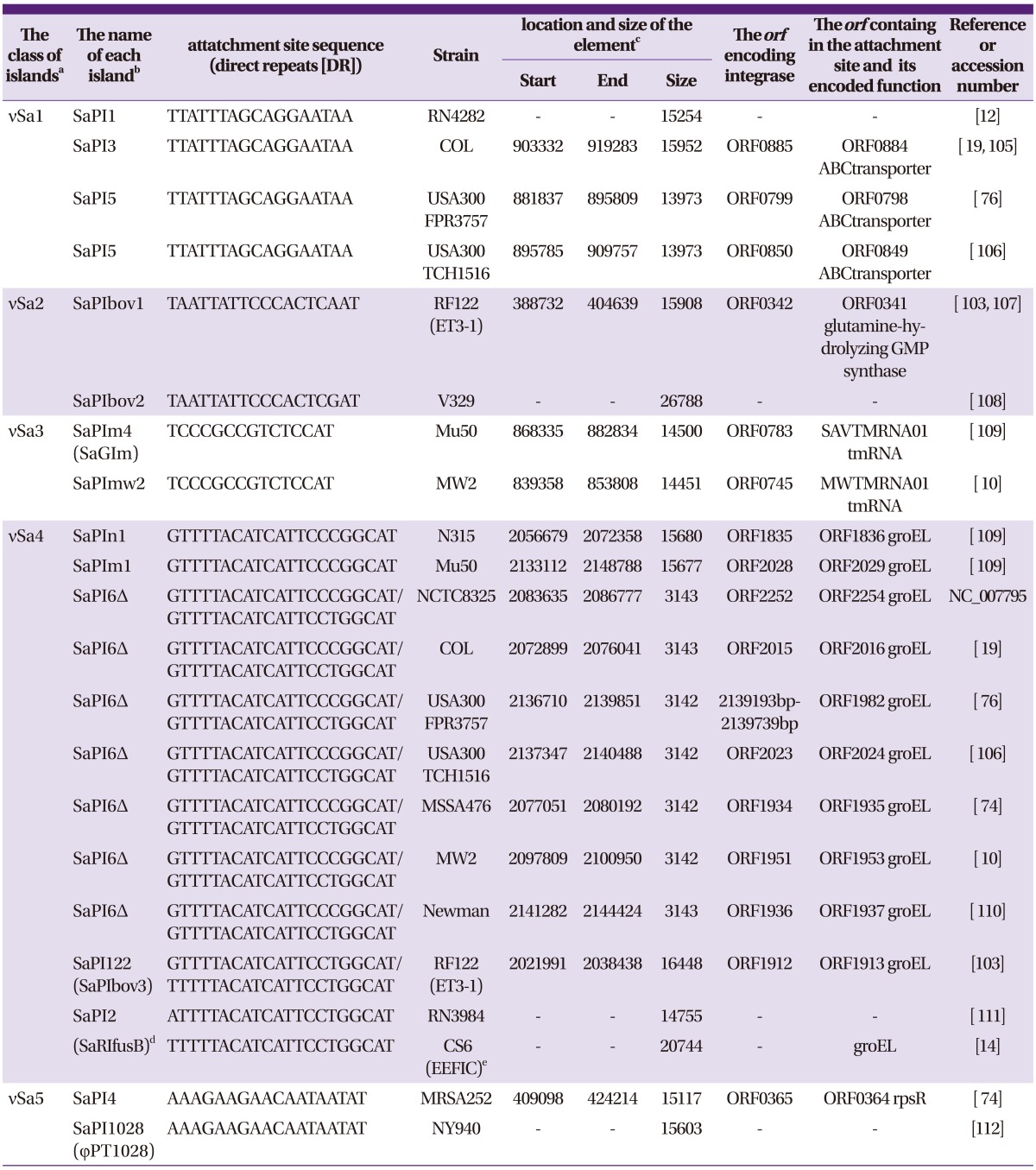
 | Figure 1
(A) Alignment of the 14 S. aureus chromosomes. Each chromosome is linearly represented with its origin of replication (oriC) placed at the left end. Using the Microbial Genome Database for Comparative Analysis [100, 101], we found 1887 orthologous open reading frames (orfs) shared by all the 14 chromosomes (denoted as grey vertical lines on each chromosome). The other orfs not commonly shared by the strains are denoted in red. The chromosomal regions containing a series of such non-orthologous orfs were extracted as genomic islands (GIs). sdh is a single gene encoding a 1.1-megadalton cell wall-associated fibronectin-binding protein [102], which is interrupted by insertion of multiple mobile elements in RF122 [103]. RF122 is the bovine isolate while the others are all human isolates. The integration site for νSaδ is present between the orfs SA2204 and SA2205 of N315. The corresponding sites are missing from the chromosomes of RF122, MSSA476, and MW2. φ signifies prophage. (B) Phylogenic tree of the integrases encoded by SaPI mobile genetic elements. The amino acid sequence homology of integrases encoded by the SaPI and SaRI mobile genetic elements listed in Table 1 was evaluated by using Clustal W WWW System provided by DNA Data Bank of Japan (DDBJ) (http://clustalw.ddbj.nig.ac.jp/top-j.html). The phylogenic tree was drawn by Phylodendron-Phylogenetic tree printer (http://iubio.bio.indiana.edu/treeapp/treeprint-sample1.html). The truncated integrases of SaPI6Δ were excluded from the analysis. Note that the phylogeny of integrase well correlates with the integraton sites of the mobile elements on the S. aureus chromosome. (See also Table 1).
|
Table 1
The list of S. aureus genomic islands
aClassified based on the integration site on the S. aureus chromosome. Correlated with the integrase phylogeny (see Figure 1B).
bThe name for the individual mobile element proposed by R. Novick [13]. Δsignifies that the element is a remnant. The name in parenthesis is the one originally described.
cStart, the first nucleoitde position of the left direct repeat (DR); End, the last nucleotide of the right DR; Size, in base pairs (bp); -, unknown because whole genome sequence data is unavailable.
dS. aureus resistance island carrying fusB [14].
eEuropean fusidic acid-resistant impetigo clone.
![]()
The SaPI elements evolved themselves to take advantage of the bacteriophage as the vehicle for efficient intercellular transmission. A representative island, SaPI1, carrying tsst-1, possesses a packaging system for phage-mediated intercellular transfer [15]. Another island, SaPIbov1, is demonstrated to be carried by a small-sized bacteriophage head that infects S. aureus cell and transfer the island into one of the several attachment sites in the chromosome [16, 17]. SaRI has essentially the same structure with SaPI, but it carries a resistance gene to fusidic acid instead of virulence gene [14].
Another class of νSa islands is found on the S. aureus chromosome. They carry many virulence-related genes, but do not have apparent device for movements. Curiously, however, a remnant of a transposase gene whose putative intact form is found in the genome of Macrococcus caseolyticus, a bacterial species sharing a common ancestor with staphylococcal species [18]. The islands νSaα, νSaβ, νSaγ, νSaδ, νSaε belong to this class of islands [10, 19], where νSaα contains a series of ssl (staphylococcal superantigen-like) genes [20, 21]; νSaδ, (previously designated νSaγ [22, 23], carries genes encoding exfoliative toxin D (ETD) and epidermal cell differentiation inhibitor (EDIN); and νSaε carries accessory gene regulators T and U involved in the virulence regulation [24], and gene clusters encoding putative lipoproteins and fibronectin-binding proteins.
Figure 2 illustrates chromosomal localization of the orfs conserved across four staphylococcal species S. aureus, S. epidermidis, S. haemolyticus and S. saprophyticus. It shows that the order of the conserved orfs (synteny) is much more retained among S. aureus strains as compared to that between the two S. epidermidis strains. This coincides with our overall impression that S. aureus has a rather conserved (or rigid) chromosome structure when compared with the other two staphylococcal species, S. epidermidis and S. haemolyticus. The chromosomes of the latter seem to be extremely labile [25, 26].
 | Figure 2
Orthologous orfs commonly shared by 17 staphylococcal strains of four species. After identifying common orthologs among various species, conserved chromosomal structure ("core structure") was constructed on the basis of the consensus arrangement of the conserved orthologs [104]. An ortholog group in the resulting core structure is indicated as a colored line across horizontal black line representing a chromosome. To simplify the figure, only universally conserved, one-to-one correspondence ortholog groups are shown. To visualize chromosomal rearrangement of the core structure, color gradation is assigned according to the location on S. aureus strain N315 chromosome from red to yellow to green. The replication origins (oriC) are located at the center. Strain-specific orfs are denoted on each chromosome by grey short vertical bars. Note that the species-specific orfs are localized in the chromosome region with sparse vertical lines.
|
The region around the origin of replication (oriC: located at the center in the illustration) is noted to be sparse for conserved orfs across staphylococcal species. This indicates that the region consists of a high proportion of species-specific genes acquired during the evolution of each species. The diversity of the chromosome is more pronounced in the region to the right of oriC (oriC to +0.5 Mb) than that to the left of oriC (-0.3 Mb to oriC). This coincides with the presence of orfX to the right of oriC, which serves as an attachment site for staphylococcal cassette chromosome (SCC), a unique horizontal gene transfer vehicle of staphylococci (discussed later). The figure also shows that the region contain a number of orfs unique to each S. aureus strain (denoted by grey short vertical bars in Figure 2) presumably acquired by SCC-mediated gene transfer. We designated this region 'oriC environ' [11]. The genetic information accumulated in oriC environ would be used by the cell to produce a more fit strain to the niche of survival.
2. Mobile genetic elements
S. aureus genome contains several copies of insertion sequences (ISs) and transposons (Tns). However, the number of copies is limited, and many of them are inactive suffering from mutations or deletions [18]. In contrast, as many as 82 IS elements are found on the S. haemolyticus genome, and most of them are intact [11]. It is curious why the number of IS or Tn are significantly different across staphylococcal species. It is well known that IS turns off a gene function by disrupting it or by inserting itself to the proximity of the target gene (polar effect) [27]. It can also up-regulate a gene transcription by integrating itself to the upstream region of a gene and providing it with a strong promoter activity [28]. In a sense, keeping many intact copies of IS and/or transposons on a chromosome is to keep the chromosome 'soft'. It is advantageous for the survival of such species of bacteria that are frequently exposed to challenging environmental conditions. The genome plasticity of S. haemolyticus is reflected in our rather frequent encounter with difficult-to-speciate clinical S. haemolyticus strains with their abnormal biochemical features compared to the standard S. haemolyticus strain [11]. Extremely high genome plasticity of S. haemolyticus might have made the species the first one to acquire vancomycin resistance [29]. It is also demonstrated that IS256 is more frequently found in nosocomial methicillin-resistant S. epidermidis (MRSE) isolates associated with septicemia and urinary tract infections as compared to commensal S. epidermidis isolates from healthy volunteers [30]. Consistent with this observation is that an oxacillin-susceptible S. epidermidis type strain ATCC12228 does not possess IS256 on the chromosome, whereas MRSE strain RP62A possesses 5 copies of IS256. Historically S. epidermidis was next only to S. haemolyticus to acquire intermediate vancomycin resistance [29]. Aside from spontaneous mutation as will be described below, IS256-mediated genome alteration might also contribute to the acquisition of vancomycin resistance in the two staphylococcal species.
In contrast to the human pathogenic staphylococci, a nonpathogenic staphylococcal species S. carnosus does not have any insertion sequences [31]. The species is isolated from meat products, and has been used safely since the 1950s as a starter culture for the fermentation of dry sausage. In the well-controlled nutrient-rich culture condition, the species would not need to alter the genome composition. So, it is reasonable that it does not maintain mobile genetic elements in the chromosome [31].
3. The oriC environ
As described above, the oriC environ is a region around the oriC where multiple exogenous genes are accumulated as a result of repeated horizontal gene transfer events. It contains a number of DNA fragments of various sizes, flanked by characteristic direct repeats (DRs). They are designated staphylococcal cassette chromosomes (SCCs) [32, 33], and are integrated in tandem in the downstream region of orfX. The DR contains integration site sequence (ISS) for SCC, which is recognized by cassette chromosome recombinase (CCR). CCR mediates integration into and precise excision of SCC from chromosome [33]. The orfX is an orf of unknown function located near oriC but well conserved among diverse gram-positive bacteria. SCC carries various exogenous genes that are considered to confer certain growth or survival advantages to the host cell. As we saw in Figure 2, the oriC environ contains many species-, and strain-specific genes. It is probable that successive integration of SCC formed the oriC environ equipped with useful genes to adapt to an unfriendly environment, and thereby contributed to the evolution of new staphylococcal strains or species.
However, the oriC environ is not a mere storehouse of foreign genes. It has another function as an extremely active diversifier of chromosome. The oriC environ serves as the generator of diversity by accumulating foreign genes, deleting useless or hazardous ones, and event inverting large chromosomal fragments across the oriC. This process is achieved by the chromosome shuffling mediated by IS and Tn copies abundantly found in oriC environ [26].
Go to :
Methicillin resistance
1. Historical accounts
Search for the genetic determinant of methicillin resistance (mec) has a long history. In 1975, Sjöström et al. demonstrated localization of mec on the MRSA chromosome [34]. In 1978 Kuhl et al. mapped mec in the linkage group II pyr-his-nov-pur of MRSA chromosome [35]. In 1980, Stewart and Rosenblum demonstrated that mec is localized on a chromosomal region (designated 'additional DNA') of MRSA, which is absent from MSSA chromosome [36]. In the early 1980s, using the penicillin-binding assay of PBP researchers noticed that a novel low-affinity penicillin-binding protein designated PBP2' or PBP2a (MecA) is produced by MRSA [37-40]. The gene mecA encoding MecA was subsequently cloned by Matsuhashi et al. by using a physically-linked tobramycin resistance determinant as a marker for cloning [41].
Matsuhashi et al. observed that mecA gene was linked to the nucleotide sequence similar to the promoter and truncated homolog of the regulator gene blaR1 of beta-lactamase (bla)-gene complex. The intact bla-gene complex has the structure blaI-blaR1-(p)-blaZ, where p stands for the divergent promoters for blaZ and blaR1. Since mecA was found in the structure ΔblaR1-(p)-mecA, they proposed the idea that mec-gene complex was produced by recombination event between mecA gene and bla-gene complex, by homologous recombination [42]. The blaI and blaR1 genes encode repressor and membrane-bound signal transducer, respectively, of blaZ transcription. Therefore, it was predictable that the integrated mecA gene in the place of blaZ was under regulatory control of the bla regulator genes. However, the strain which Matsuhashi used for molecular cloning did not contain an intact set of regulator genes. Tesch et al. was the first demonstrating the presence of the region (called mecR locus) upstream of mecA of methicillin-resistant S. epidermidis strain WT55 that strongly repressed expression of MecA [43].
There were methicillin-resistant strains with and without the mecR locus [44, 45]. Those having the mecR locus tend to be slow in the induced production of MecA by exposure to methicillin [46]. We also identified the mecR locus in S. aureus strain N315 that was curiously methicillin-susceptibile irrespective of the presence of mecA gene. The mecI-mecR1 regulator genes homologous to blaI-blaR1 were found in the mecR locus of N315 [47]. Inactivation of mecI gene de-repressed production of MecA, and induced expression of methicillin resistance [48]. The mecA gene transcription is also repressed by bla-gene complex on a plasmid harbored by N315, but the bla regulators allowed much quicker induction of mecA on exposure to methicillin. As expected, mecI gene was either deleted or mutated in most of the clinical MRSA isolates having apparent methicillin-resistant phenotype [1]. There were also some strains with intact mecI gene but having a mutation in the operator region of mecA gene to which MecI repressor protein is supposed to bind [1]. The mecA-carrying methicillin-susceptible strains as N315 are rarely found in clinical samples. It is consistent with the observation that N315 generates mecI-inactivated mutants at high frequencies upon beta-lacatm selection in vitro. Such strain as N315 having depressed MecA expression due to the intact mecI was considered as a precursor of MRSA, designated pre-MRSA [1, 48].
2. SCCmec as a vehicle of methicillin resistance determinant
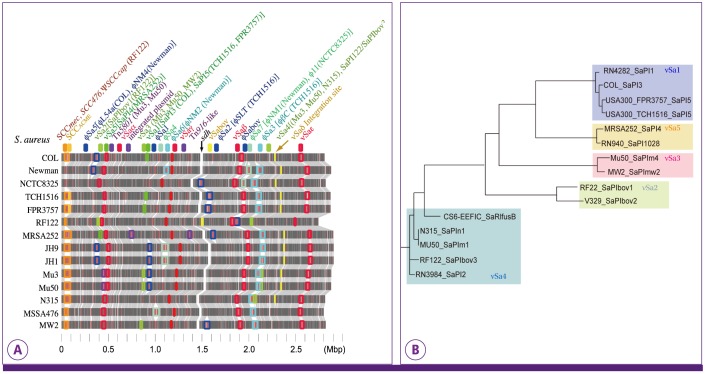
Using three epidemiologically representative MRSA strains with different genetic backgrounds, we determined the nucleotide sequence of entire additional DNAs [1, 49]. They appeared as long (32->60 Kb) stretches of chromosomal DNA regions with many orfs of unknown function within which two important clusters of genes were identified; mec-gene complex (encoding methicillin resistance) and cassette chromosome recombinase (ccr) -gene complex (encoding one or two site-specific recombinases for the movement of the element) [50] (Fig. 3A). These long DNA regions were found integrated site-specifically at the integration site sequence (ISS) present at the 3' end of orfX as if it were a 'cassette' DNA. We designated the element staphylococcal cassette chromosome mec (SCCmec) [33]. However, unlike the refined image of the 'cassette' for integron [51], SCCmec elements were much larger and studded with apparently useless pseudogenes or truncated copies of transposons and insertion sequences. This is the reason why the authors called the regions around the two essential gene clusters as J (standing for 'junkyard') regions [52]. Now, the J region is spelt out as 'joining' and its sequence is used for the subtyping of various SCCmec types [53, 54].
 | Figure 3
(A) Basic structure of SCCmec. SCCmec is bracketed by direct repeats (DRs) that contain integration site sequence (ISS) recognized by cassette chromosome recombinase (CCR). A pair of inverted repeats (IRs) are present at the termini of SCCmec. Two critical gene complexes, ccr and mec are present, and the other regions are designated J1, J2, and J3. The type of SCCmec is defined by the combination of the type of ccr-gene complex and the class of mec-gene complex. Subtype of the SCCmec is based on the difference in the J (standing for junkyard) regions. (B) Various types of SCCmec. The structures of 11 types of SCCmec are illustrated based on the nucleotide sequences deposited in the DDBJ/EMBL/GenBank databases as follows: type I, NCTC10442 (AB033763); type II, N315 (D86934); type III, 85/2082 (AB037671); type IV, CA05 (AB063172); type V, WIS [WBG8318] (AB121219); type VI, HDE288 (AF411935); type VII, JCSC6082 (AB373032); type VIII, C10682 (FJ390057); type IX, JCSC6943 (AB505628); type X, JCSC6945 (AB505630), and type XI, LGA251 (FR821779.1) Direct repeats that comprise integration site sequences of SCC are located at both extremities of SCCmec (the red arrowheads). The location of five (A-E) classes of mec-gene complexes is indicated by pink belt. The locations of ccr-gene complexes are indicated by blue belt. Insertion sequences and transposons are indicated in yellow. Representative genes related to heavy metal resistance and integrated plasmids located in the J regions are also indicated. Type XI is a newly identified SCCmec found in the MRSA strains of bovine sources.
|
As illustrated in Figure 3A, SCCmec is demarcated by specific inverted repeats (IR) and direct repeats (DR) that contain integration site sequence acted on by cassette chromosome recombinase (CCR). The element carries the mecA gene, as a component of mec-gene complex, and ccr gene(s) (ccrAB or ccrC) that encode CCR(s). The integration site for the cassette chromosome is present at the 3'-end of an open reading frame, orfX. As a result of integration, the SCC element is flanked by a couple of direct repeat (DR) sequences, both of which serve as integration sites for the next SCC element (Fig. 3A). The function of orfX is unknown, but it may encode an important protein. Upon integration of SCCmec, 15 bp of the 3'-end of orfX are replaced by the terminal nucleotide sequence of the integrated SCCmec. Curiously, however, in spite of the difference in the nucleotide sequence, the encoded peptide is unchanged before and after the integration of any type of SCCmec [49, 55, 56].
Many types of SCCmec have been found (Fig. 3B). At present, 11 types are registered [57], and more numbers are expected to be found in the future. The SCCmec type is defined by the combination of the type of ccr-gene complex and the class of mec-gene complex. Types I-III are older SCCmec types that are harbored by HA-MRSA [49]. They are relatively big in size and carry multiple antibiotic resistance determinants. Types IV and V were recognized as new versions of SCCmec that were almost diagnostically harbored by CA-MRSA [55, 56, 58]. They are short, and typically carrying no antibiotic resistance genes other than mec-gene complex. These new types of SCCmec are widely distributed among methicillin-resistant non-aureus staphylococci (MRNAS) especially among methicillin-resistant S. epidermidis (MRSE) carried by Japanese healthy children in the community [25, 59]. Recent study performed in other countries showed wide distribution of type-IV SCCmec among MRSE and type-V SCCmec in methicillin-resistant S. haemolyticus (MRSH) in the community [60]. These data indicate that MRNAS serves as the reservoir for horizontal gene transfer of new types of SCCmec to S. aureus.
3. New types of SCCmec: cost of resistance and influence on virulence
Integration of SCCmec into the oriC environ of MSSA chromosome generates MRSA. Therefore, each clone of MRSA (clonotype) can be described by the combination of the genotype of recipient MSSA strain and the genotype of the integrated SCCmec [3, 50, 61]. Recent worldwide outbreak of community-acquired MRSA (CA-MRSA) is caused by the emergence of new molecular types of SCCmec [58]. MRSA carrying new types of SCCmec (types IV and V) appeared in the last decade and almost simultaneously from various countries in the world [55,56]. As shown in Figure 3B, types IV and V are discriminated from the previously dominant three types of SCCmec (mostly carried by HA-MRSA) with different combinations of mec-gene complex and ccr-gene complex. When compared with HA-MRSA strains with type I-III SCCmec, CA-MRSA strains carrying type-IV and type-V SCCmec tend to be faster in growth, more virulent, resistant to fewer antibiotic classes (non-multi-resistant), and lower in the degree of beta-lactam resistance (heterogeneously methicillin resistant) [58]. These features of CA-MRSA strains seem to be associated with the structural feature of the new types of SCCmec. The most evident first feature is the small size of the type -IV and -V SCCmec: they carry less number of exogenous genes that might disturb physiology of the cell, and no antibiotic resistance gene against various antibiotic classes besides mecA gene [58]. The smaller size of the elements might have helped them distribute in such an explosive way to community S. aureus strains by efficient horizontal gene transfer presumably via bacteriophages [62].
Another feature is observed with type-IVa SCCmec, which is widely disseminated in CA-MRSA [58]. All the eight type-IVa SCCmec elements so far sequenced share a mutation in the Shine-Dalgarno (S-D) sequences for mecA gene (they have -AGGAGT- instead of -AGGAGG-). This might explain the low phenotypic expression of methicillin resistance (or hetero-methicillin resistance) with oxacillin MIC values of 4-64 mg/L of the CA-MRSA strains carrying the type of SCCmec [58]. In most of the MRSA strains, the production of MecA encoded by mecA gene is repressed by BlaI repressor protein encoded by blaI gene on the penicillinase plasmid [63]. De-repressed expression of the exogenously acquired cell-wall synthesis enzyme MecA seems to cause disadvantage to the cell presumably by disturbing otherwise well organized cell-wall synthesis performed by multiple intrinsic enzymes. In fact, removal of penicillinase plasmid de-represses mecA gene expression and causes frequent loss of mecA gene during drug-free passage [64, 65]. Lee et al. also examined the cost of acquisition of SCCmec using an MSSA strain NCTC8325 (BB255) [66]. The doubling time of the MSSA strain was 39.3 min. The doubling time of the transformant with type-I SCCmec increased significantly to 54.5 min, whereas that with the type-IV SCCmec was 41.3 min. Moreover, type-IV SCCmec did not impose an energetic cost to its recipient MSSA with regard to the amount of the cell mass produced by consuming a unit of ATP. On the other hand, the presence of type-I SCCmec significantly reduced the cell yield [66]. Type-I SCCmec appears to have a negative effect on the growth rate and the effective cell reproduction compared to type-IV SCCmec in batch culture. The reason for this difference is yet to be examined, however, Lee et al. referred to the possible contribution of the mutated S-D sequence for mecA gene in the type-IV SCCmec used for the experiments [66]. Low expression of MecA due to the mutated S-D sequence may make the cell more fit in the survival in the community where exposure to beta-lactam antibiotics is less frequent than in the hospital.
The third intriguing feature of new SCCmec is the absence of psm-mec gene encoding phenol-soluble modulin [67]. It is present adjacent to the mecI gene of class-A mec-gene complex (Fig. 4). Although S. aureus is non-flagellated and unable to move, but some strains as MSSA strain Newman tend to spread on soft agar forming diffuse colonies [68]. Introduction of the psm-mec found in class-A mec gene complex into Newman suppressed its spreading ability and decreased exotoxin production. Introduction of psm-mec also attenuated the virulence of Newman in a murine systemic infection model [69]. Presence of psm-mec decreases the production of cytolysin PSMα. PSMα is another phenol-soluble modulin that augments cytokilling activity of Panton-Valentine leukocidin [70]. Both the transcription and translation products of the psm-mec suppressed colony-spreading activity and promoted biofilm formation [71]. This psm-mec RNA was found to bind agr mRNA and inhibit translation of the agrA gene [72]. Since agrA encodes a positive transcription factor for the psmα gene, the production of PSMα is depressed. The gene is found immediate downstream of mecI gene of type-II, type-III, and type-VIII SCCmec elements, whereas it is absent from type-IV and V SCCmec elements [69]. The absence of the gene from the new types of SCCmec explain at least partially the higher virulence of CA-MRSA strains as compared to HA-MRSA.
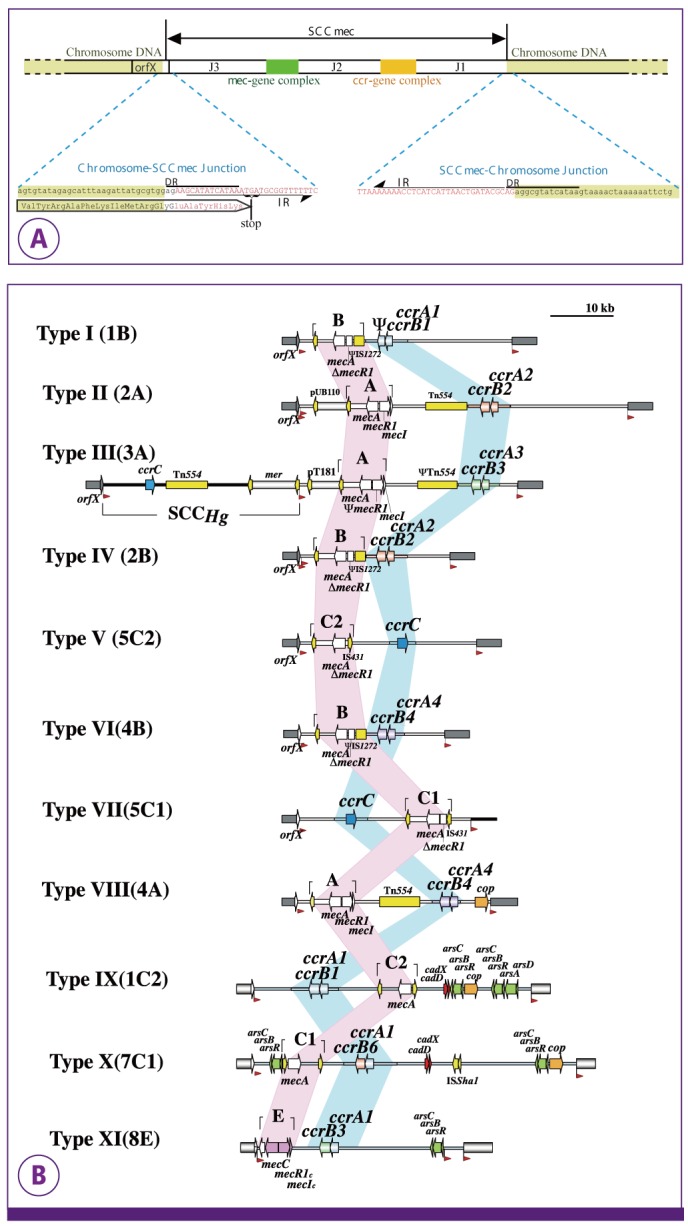
 | Figure 4The mecA gene originated from Staphylococcus fleurettii genome.
Practically identical mecA gene and its surrounding region of SCCmec were identified on the chromosomes of S. fleurettii, a member of the sciuri group of staphylococcal species. The mecA-gene orthologs found in other staphylococcal species at the corresponding chromosomal loci (mvaS, a vital gene for life, serves as a landmark for the locus) are either deleted or degenerated presumably due to disuse of their function. Two mobile elements IS431 and Tn554 seem to have excised this part of the chromosome and inserted it into an SCC to form the historically first SCCmec element.
|
4. SCC as a vehicle for genes other than mecA
When SCCmec was found, it was considered that it is an exclusive vehicle for mec-gene complex and some other drug-resistance genes (Fig. 3B). Subsequently, however, SCCs harboring other functional genes than mecA started to be found in the oriC environ. They carried type-1 capsule gene cluster (SCCcap1) [73], fusidic acid resistance gene (SCCfur) [74], and mercury resistance operon (SCCHg) [53], penicillin-binding protein 4 [75], and enterotoxin H (seh) [10]. Recently, an SCC-like element carrying an arginine deiminase pathway and an oligopeptide permease system (designated arginine catabolic mobile element [ACME]) was found next to a type-IV SCCmec element in a CA-MRSA strain prevailing in the USA (designated USA300) [76]. The element is considered to confer better survival ability and increased virulence to the strain. With these observations, now it seems likely that SCC serves as a vehicle for not only drug resistance genes but also diverse genes useful for better adaptation to the unfriendly environment. Multiple SCC elements can be integrated sequentially into one of the DRs bracketing the previously integrated SCC elements. Thereby, accumulation of multiple SCC elements carrying various functions is achieved to allow diversification of S. aureus genome.
5. Origin of mecA gene
Search for the origin of mecA gene was an important project for the researchers of MRSA. First, a mecA gene homolog encoding a PBP with 88% amino acid similarity to MecA was found on the chromosome of S. sciuri strains [77]. Then a mecA gene homolog with practically identical nucleotide sequence (greater than 99%) with mecA was discovered on the chromsome of S. fleurettii [78], one of the oldest staphylococcal species. As shown in Figure 4, the chromosomal locus containing the complete mecA-gene complex as well as the surrounding J regions bracketed by two mobile genetic elements Tn554 and IS431 of type-II SCCmec was found in S. fleurettii strain SFMP07 [78]. Subsequent whole genome sequencing of the strain revealed that the region was about 240-Kb apart from the orfX, and there were multiple SCCs in the vicinity of orfX but no copy of it found around the mecA gene complex (Tsubakishita et al. in preparation). This indicated that mecA was an intrinsic component of the chromosome of S. fleurettii. It must have been an important PBP for the ancestral staphylococci to survive the environment soaked with ß-lactam antibiotics produced by fungi and Actinobacteria. The mecA gene must have been transmitted vertically during the initial steps of staphylococcal speciation. However, after a while, it became decayed (with mutations) or deleted out from the chromosomes of the descendants as exemplified by the emergence of methicillin-susceptible S. aureus (MSSA) as a human colonizer. It is curious why mecA gene was lost from the staphylococcal chromosome during speciation. The divergence time of S. sciuri group from the major staphylococcal clade including S. aureus is calculated to be 200-300 million years ago [79], which corresponds to the geological age of the emergence of mammalian animals. Since then, staphylococcal species evolved with mammalians as the colonizers of diverse mammalian animals [79]. Our hypothesis is that the descendants of staphylococci became methicillin-susceptible, because they became protected from the threat of ß-lactam-producing fungi or actinobacteria by the immune system of mammalian hosts [79].
6. mecB and mecC, new mecA gene homologs
SCC is a mobile genetic element found only in staphylococcal species and in macrococcal species that are close relatives of the former. The two genera share the same ancestor and diverged from each other about 300 million years ago as inferred by the phylogenetic tree with 16S and 23S DNA sequences and calculation based on the ecological distribution study of staphylococci across mammalian animals [79]. Genetic study on several ß-lactam-resistant Macrococcus caseolyticus strains isolated from China, Japan, and Thailand, revealed that they did not harbor mecA. Instead, a novel PBP designated mecB was identified on a plasmid, which turned out to be a distantly related mecA-gene homolog, and was shown to confer methcillin resistance on the cell. The gene had 61.7% of nucleotide identity with mecA (Fig. 5). The mecB was also accompanied by regulator genes mecR1b and mecIb that were also distantly related to the mecR1a (53.5% identity) and mecIa (60.2% identity), respectively, of the mecA-gene complex. Curiously, however, unlike mecA gene, mecB was accompanied by a blaZ homolog (blaZb) encoding a putative beta-lactamase. Moreover, unlike mecA-gene complex, the mecB-gene complex (blaZb-mecB-mecR1b-mecIb) was adjoined to transposase genes, forming transposon Tn6045 (Fig. 5). The mecB transposon was found on either plasmid (Fig. 6) or chromosome of macrococcal strains. One strain JCSC7096 harbored the transposon on the chromosome next to orfX, and an SCC element carrying ccrAB genes was found right next to the transposon [80]. Only one copy of the direct repeats of the transposon separated the two elements, Tn6045 and the SCC. A mere mutational inactivation of transposase or deletion of the direct repeat would be enough to combine the two elements into a new SCCmec [80]. The new SCCmec may come into the strains of staphylococci in near future, or might have already been there to be found by the researchers.
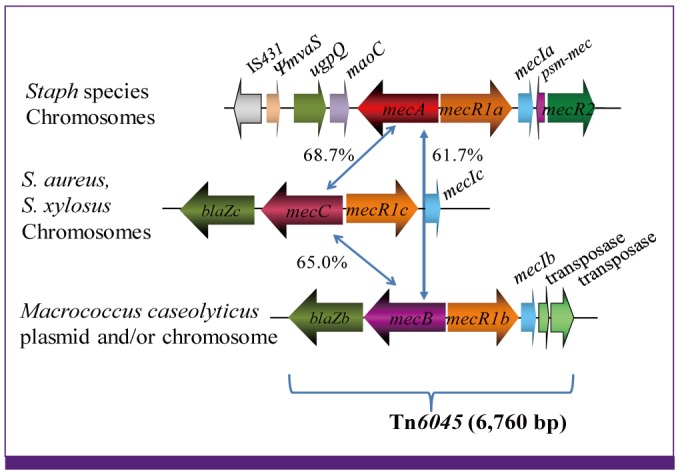 | Figure 5Novel mecA gene homologs mecB and mecC.
mecB (formerly mecAm) and mecC, are found linked with blaZ homolog encoding beta-lactamase. mecB-gene complex is associated with transposase genes and transmitted as transposon Tn6045. mecA- and mecC-gene complexes are transmitted as SCCmec elements.
|
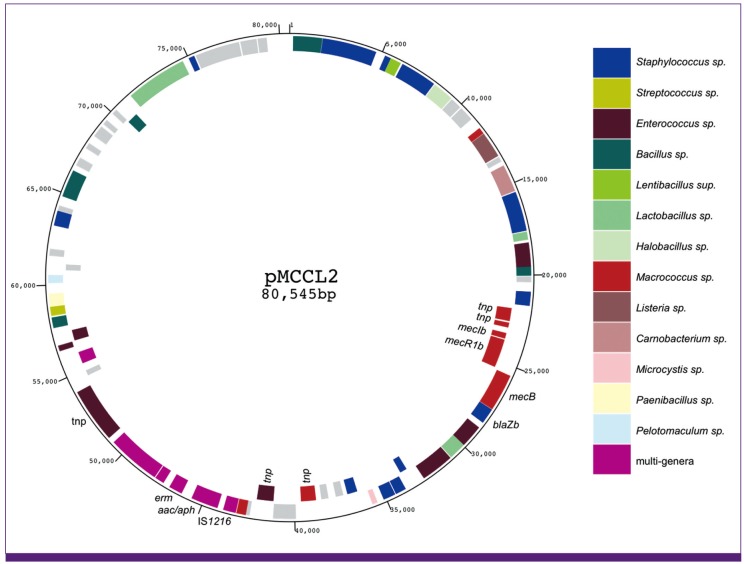 | Figure 6
mecB transposon Tn6045 is carried by a big R plasmid.
Physical map of the Macrococcus caseolyticus plasmid pMCCL2 is illustrated [18]. BlastP analysis was performed on the protein product of each orf. The bacterial genus of the top-hit entry for each orf is denoted by coloration. Those having no entry with e-value of 1 × 10-6 or above, the orfs are colored in grey. The plasmid contain antibiotic resistance genes such as erm and aac/aph genes besides mecB transposon. Note that the plasmid contains orfs whose top-hit entry convers as many as 13 different genera.
|
Recently, another mecA-gene homolog designated mecC was found on the chromosome of MRSA strains of animal as well as human sources [81, 82]. mecC was almost equally distant from mecA and mecB, and shared the characteristic gene order of blaZc-mecC-mecR1c-mecIc, having a blaZ-gene homolog next to mecC (Fig. 5). However, unlike mecB-gene complex, the mecC-gene complex was not associated with transposase genes. Instead, it was linked to ccr-gene complex forming a novel SCCmec (type XI; see Figure 3B). So far, mecC has also been found on the chromosome of S. xylosus [83], but mecB has yet to be found in staphylococcal species. A possible explanation for this difference might be due to the host specificity of the mobile genetic elements that carry them; mecA and mecC are carried by SCC, and mecB, by transposon. However, the plasmid carrying the mecB transposon seems to have a broad host range (Fig. 6). It is a resistance (R) plasmid carrying genes with diverse blast top-hit entries across several other bacterial genera besides Macrococcus and Staphylococcus. They cover more than 10 genera including Enterococcus, Bacillus, and Streptococcus (Fig. 6). It is speculated that mecB is provided to diverse bacterial genera via the plasmid. And yet, mecB has not been found in staphylococcal species. SCC may be a more efficient genetic transfer machinery than plasmid or transposon across staphylococcal species, although we do not know its inter-species transfer mechanism. It would be an important project to look for the vehicle of SCC transmission to reveal seemingly an efficient genetic information exchange system of staphylococcal species.
7. Mec superfamily
Figure 7 shows the evolutionary tree of penicillin-binding proteins (PBPs) of Staphylococcus species. S. aureus has four PBPs, and MRSA has MecA or MecC acquired by horizontal transfer as the fifth PBP. When the Macrococcus MecB was used for the homology search, other genera such as Enterococcus species were found to have mec gene homologs. The pbp homologs possessed by Macrococcus caseolyticus came between those of the enterococci and staphylococci in terms of the degree of homology. The MecA homologs with 83-90% amino acid (aa) identities were found in non-aureus staphylococcal species [77, 84]. With the discovery of the Macrococcus MecB having 52% aa identity to MecA, all the Mec homologs including recently identified MecC were grouped as members of the Mec superfamily together with a group of enterococcal PBPs (Fig. 7). This is a significant observation, because it has long been known that enterococcal species especially E. faecium is naturally resistant to beta-lactam antibiotics [85]. The degree of resistance as evaluated by beta-lactam MIC was well correlated with the degree of binding saturation of PBPs produced by enterococcal strains [86]. Thus the binding affinity of the PBPs and the amount of their production are considered to determine the level of susceptibility of the strain towards beta-lactam antibiotics. PBP5 of E. faecium was shown to have the lowest binding affinity to beta-lactam antibiotics. Similarity between MecA and enterococcal low-affinity PBPs was pointed out when mecA gene was cloned and sequenced [87]. By the way, a pbp gene was found on a plasmid harbored by a beta-lactam resistant Enterococcus hirae strain. The predicted protein encoded by the gene, designated PBP3r, had 99.7% amino acid identity to PBP5 of E. faecium [88]. Therefore, it seems likely that not only MecB but also other ß-lactam-insensitive pbp genes were valuable trade articles among bacterial species annoyed by ß-lactam-producing fungi or actinobacteria.
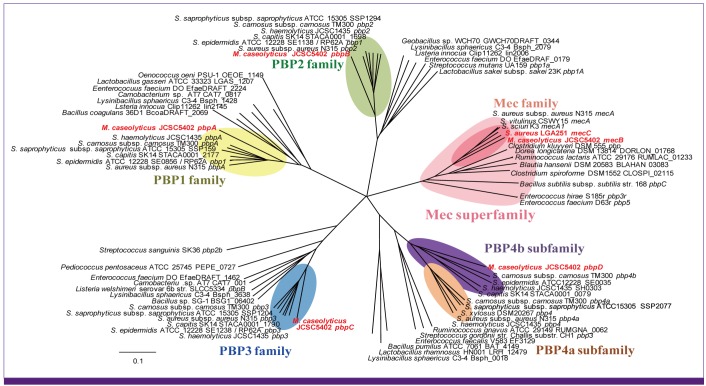 | Figure 7Phylogenic tree of Penicillin-Binding Protein (PBP) genes homologous to the Macrococcus caseolyticus PBP genes.
M. caseolyticus carries four PBP genes, pbpA-D on the chromosome and mecB on the plasmid pMCCL2. BlastX was performed with each of the five PBP genes as query. The genes with lower than 55% similarity or 50% query coverage to each of the PBP genes of M. caseolyticus were declined for subsequent analysis. All the used nucleotide sequences were obtained from GenBank database (http://www.ncbi.nlm.nih.gov/Genbank/). The phylogenetic tree was generated using the neighbor joining method in the ClustalW program (http://clustalw.ddbj.nig.ac.jp/top-j.html). The tree was visualized in the Phylodendron Web Form (http://iubio.bio.indiana.edu/treeapp/treeprint-form.html). To simplify the tree, we adopted the pbp homologs of strain N315 as the representatives of S. aureus pbps, all the pbp homologs of non-aureus staphylococcal species, and the most homologous pbp from each of the non-staphylococcal bacterial families. Macrococcus genes are depicted in red. Note that pbp1-3 genes of various gram-positive bacteria are ordered in a similar phylogenetic relationship, whereas the entry of bacterial families or species carrying mecA and pbp4 homologs and their relative phylogenetic distance are significantly different from the former. This indicates that mecA and pbp4 are laterally transmissible genes.
|
8. PBP4 as another mobile PBP
Figure 7 also reveals a curious fact that PBP4 is classified into two subgroups based on the Blast homology analysis. One group is named PBP4a that contains S. aureus PBP4, and the other, PBP4b to which Macrococcus PBP4 belongs (Fig. 7). Figure 8 shows the relative chromosomal locations and direction of transcription of the pbp genes on the macrococcal and staphylococcal chromosomes. The relative locations for pbps1-3 are well conserved among M. caseolyticus and four staphylococcal species. The location of pbp4a relative to pbps1-3 is conserved in several staphylococcal species. However, no pbp4 is found in S. epidermidis strain RP62A, and pbp4a is absent but pbp4b is present in another S. epidermids strain ATCC12228. Curiously, both pbp4 subtypes are present in S. haemolyticus strain JCSC1435 and S. carnosus strain TM300. It is also remarkable that, like mecA gene, most of the pbp4 genes are localized in the oriC environ (Fig. 8). The pbp4b of S. epidermidis strain ATCC12228 in Figure 8 was actually carried by an SCC element designated SCCpbp4 [89]. Therefore, it is likely that at least a part of pbp4 genes on staphylococcal species were acquired by horizontal gene transfer, whereas the set of three pbps1-3 were vertically transmitted from the common ancestor of macrococcal and staphylococcal species.
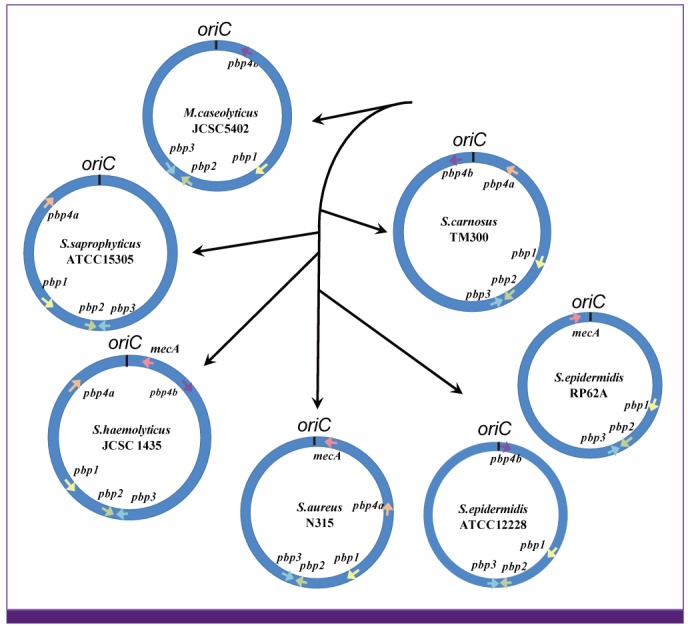 | Figure 8Relative position of pbp gene homologs on the chromosome of staphylococcal species.
The figure shows relative location of the pbp genes on each chromosome. The arrow on the chromosome denotes the pbp gene and its direction of transcription. The relative placement of the chromosomes and black arrows represent a rough phylogenic tree of staphylococcal species based on 16S ribosome sequence. The distance between each species is not to the scale. Only the order of branching (speciation) from ancestral bacteria is shown. The relative locations and directions (synteny) of pbp1-3 genes are well conserved among staphylococcal species. Note that synteny is incomplete with pbp4 and mecA. They are located around the oriC (the oriC environ). In S. carnosus, two types of pbp4 genes, denoted as pbp4a and pbp4b, are noticed [31].
|
In the case of S. epidermidis strains, vertically transmitted pbp4a might have been lost from RP62A by chromosome shuffling in the oriC environ, and ATCC12228 acquired pbp4b homolog as an SCC element by horizontal gene transfer. PBP4 is not essential for the viability of S. aureus [90], but its overproduction raises resistance to beta-lactam antibiotics [91]. By functioning as a transpeptidase, PBP4 increases peptidoglycan cross linkage, giving the cell wall a more rigid structure. Therefore, like mecA, pbp4 seems to serve as a mobile pbp to aid ß-lactam resistance of staphylococcal and related bacterial species.
9. Phenotypic expression of methicillin resistance
Acquisition and expression of mecA gene alone do not make the cell uniformly resistant to ß-lactam antibiotics. The expression of MecA confers on the bacterial strain a moderate level of resistance to ß-lactam antibiotics. The strain shows a unique pattern of antibiotic resistance called 'hetero-resistance'. Such MRSA is called hetero-MRSA. Figure 9 shows a typical ß-lactam (methicillin)-resistance profile of a hetero-MRSA (strain N315ΔIP) to which 64 mg/L of methicillin is effective to suppress the growth of 99.9% of N315ΔIP (Fig. 9). However, the rest of the cell population does grow. The population is composed of heterogeneous subpopulations of cells with higher methicillin resistance in different degrees. Figure 9 shows that a minor subpopulation of N315ΔIP consisting about one in 1 million part of the entire cell population can grow even in the presence of 1,024 mg/L of methicillin (Fig. 9). Exposure of the hetero-MRSA strain to a potent ß-lactam such as imipenem easily selects out a mutant strain, whose entire cell population is uniformly highly resistant to ß-lactam antibiotics, called homogeneously methicillin-resistant S. aureus (homo-MRSA) [61, 92]. This change, designated 'hetero-to-homo conversion' of ß-lactam resistance, is achieved by spontaneous mutations on the chromosome designated chr* [93].
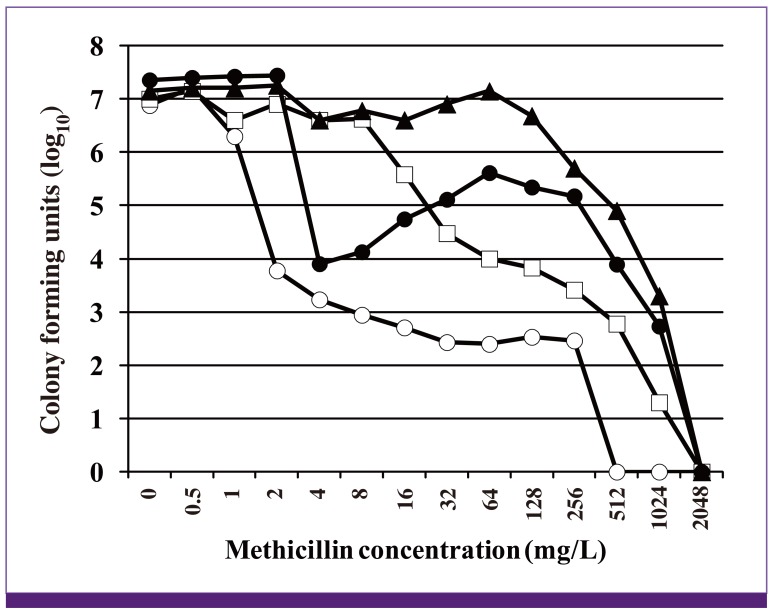 | Figure 9Four phenotype expressions of methicillin resistance.
Symbols: open circle, pre-MRSA strain N315 (mecA gene is only weakly induced in N315, because it is strongly repressed by the repressor protein encoded by an intact copy of mecI gene. Spontaneous mutational inactivation of mecI gene makes the cell express methicillin resistance by allowing constitutive production of the mecA gene transcript [48]); open square, hetero-MRSA strain N315ΔIP (mecI gene-deletant of N315 expressing hetero-methicilin resistance); closed triangle, homo-MRSA strain N315ΔIP-H5 (obtained by selecting N315ΔIP with 8 mg/L of imipenem. Single mutation rpoB(N967I) is responsible for the 'hetero-to-homo conversion'); closed circle, strain N315rpoB(N967I) (the chr* mutation rpoB(N967I) was introduced into N315 by gene replacement procedure). Note that chr* causing hetero-to-homo conversion confers 'Eagle-phenotype' on the pre-MRSA [97].
|
So far mutations in vraSR (standing for vancomycin resistance sensor and response regulator) [94, 95], a two-component regulator system for S. aureus cell-wall synthesis, have been identified as chr*. Mutations vraS (H5N) and vraS (S329L) constitutively activate the regulatory system, and enhance cell-wall synthesis of the cell. Concomitant increase in resistance was observed against cell-wall synthesis inhibitor antibiotics such as ß-lactam, fosfomycin, bacitracin, and glycopeptides [94].
10. Regulatory mutation for the expression of methicillin resistance
Now, extensive search for chr* can be done using high thorough-put whole genome sequencing strategy. By comparing the genome of hetero-MRSA strain ΔIP and its derivative strain ΔIP-H5, a mutant strain obtained by selection of ΔIP with imipenem, we found a mutation rpoB (N967I) as a chr* for the hetero-to-homo conversion of MRSA [96]. Likewise, another mutation rpoB (R644H) was identified from a N315-derived strain N315h4 expressing 'Eagle-type' methicillin resistance [97]. Eagle type resistance has a unique population analysis curve as illustrated in Figure 9: more cells grow in the presence of higher concentrations of methicillin (32-256 mg/L) than in low concentrations (4 and 8 mg/L). Eagle-type resistance is known to be a special phenotypic expression of chr* in the 'pre-MRSA' strain N315 in which mecA gene expression is strongly repressed under the repressor function of mecI gene [48]. Since induction of mecA gene transcription by low concentrations of methicillin is weak and slow in pre-MRSA, cells die. The rpoB mutations seem to confer on the cell methicillin 'tolerance', and allow the cell survive in methicillin until enough amount of MecA is accumulated in the cell [48, 96].
rpoB gene encodes RNA polymerase ß subunit RpoB. Mutation of rpoB has long been assigned significance only as the resistance marker for rifampin. Recently however, we started to notice that rpoB mutations affect various antibiotics other than rifampin (Fig. 10). One of the most frequent mutations rpoB(H481Y) raises not only rifampin resistance but also vancomycin-intermediate resistance of S. aureus [98]. Another mutation rpoB (Q468K) raises resistance to both rifampin and daptomycin. Mutations rpoB (T480M) and rpoB (R503H) do not affect rifampin susceptibility, and yet raise resistance to teicoplanin, vancomycin, and daptomycin [98]. The two mutations rpoB (N967I) and rpoB (R644H) do not affect susceptibility to rifampin either, but cause hetero-to-homo conversion of methicillin resistance (Fig. 10) [96]. Curiously, practically all rpoB mutations so far identified conferred enhanced susceptibility to linezolid [98].
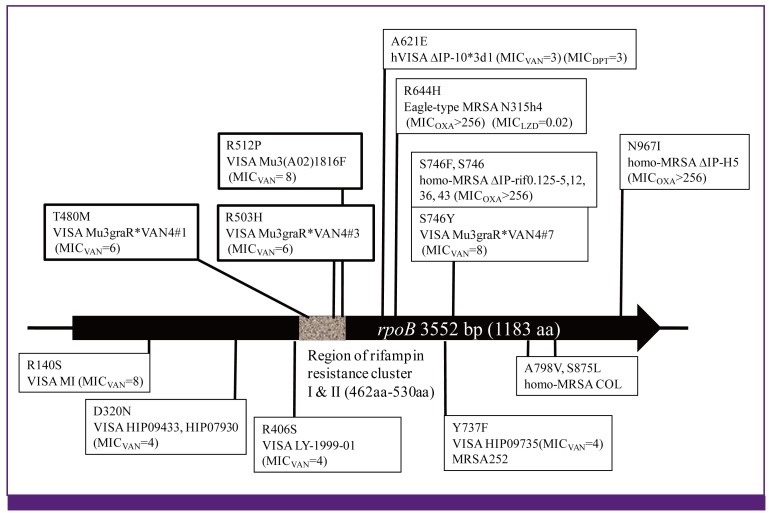 | Figure 10
rpoB gene mutation as the 'regulatory mutation'.
Those rpoB gene mutations found in-vitro-derived mutant strains and the phenotype of the strains are listed above the rpoB gene figure. Those in clinical strains that express altered susceptibilities to vancomycin, ß-lactam, daptomycin and linezolid, but susceptible to rifampin (MICRIF < 1.0 mg/L) are listed under the rpoB figure. The mutations in the clinical strains remain to be demonstrated for their direct contribution to the phenotypes.
|
Depending on the locations of the mutations in rpoB gene and the kinds of amino acid substitutions the repertoire of affected antibiotics and the degrees of resistance are different [98]. Matsuo et al. showed that the resistant subpopulations of heterogeneously vancomycin-intermediate S. aureus (hVISA) contain the cells with many different rpoB mutations [98]. hVISA and hetero-MRSA share a similar population analysis curve. Therefore, it is likely that rpoB mutations are one of the major classes of mutations that determine the characteristic shape of the population curve of both hetero-MRSA and hVISA (Fig. 9).
rpoB mutations are well known to cause rifampin resistance by changing the conformation of the rifampin-binding site of RpoB. However, there are another class of rpoB mutations that decrease susceptibility to such antibiotics as glycopeptides, ß-lactam, and daptomycin without affecting rifampin susceptibility. RpoB is not the target of these antibiotics. Therefore, those rpoB mutations are considered to change the antibiotic susceptibility of the cell indirectly by changing the physiology of the cell. In this sense, those rpoB mutations are regarded as 'regulatory mutations'. They confer changes in transcription profile of the cell by altering the interaction preferences of RNA polymerase holoenzyme for diverse transcription regulator proteins.
Go to :
Future perspective
Now, with a great technical advance in high thorough-put determination of nucleotide sequence, it became easy to determine and compare hundreds of bacterial genomes. By doing this, we will obtain a bird's view of the evolution of life on the earth. Antibiotic chemotherapy has long been faced with a difficult problem of antibiotic resistance. It is now evident that no antibiotic can escape the doom of emergence of resistance. As we have seen, however, methicillin resistance is likely to have existed long before the advent of mammalians in the history of life. We may reasonably think that antibiotic resistance is not an outcome of human misconduct. Apparent vicious cycle of antibiotic and antibiotic resistance may be regarded as a providence of nature. We recently found a curious class of antibiotic designated Reverse Antibiotic (RA), which was active against quinolone-resistant S. aureus but inactive against quinolone-susceptible S. aureus [99]. RA was found in the soil of earth as a natural product. It is futile to try to disrupt the vicious cycle by incessantly producing new antibiotics. Instead, we may learn how to control the procession of the vicious cycle by closely observing the life history of earth, where all the opponent microorganisms seem to coexist in a dynamic equilibirium.
Go to :
 XML Download
XML Download