

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
It was previously observed that recombinant flock house virus (FHV) RNA1 was efficiently packaged into turnip yellow mosaic virus (TYMV), provided that the TYMV coat protein (CP) sequence was present at the 3′-end. FHV RNA encapsidated by TYMV CPs also had a four-nucleotide extension at the 5′-end. Since even a short extension at the 5′- and 3′-ends of FHV RNA1 inhibits replication, we examined whether the recombinant FHV RNA is indeed capable of replication. To this end, we introduced constructs expressing recombinant FHV RNAs into the plant Nicotiana benthamiana. Northern blot analysis of inoculated leaves suggested abundant production of recombinant FHV RNA1 and its subgenomic RNA. This demonstrated that recombinant FHV RNA with terminal extensions at both ends was competent for replication. We also showed that the recombinant FHV RNA can express the reporter gene encoding enhanced green fluorescent protein.
Go to : 
REFERENCES
1). Geall AJ, Mandl CW, Ulmer JB. RNA: The new revolu-tion in nucleic acid vaccines. Semin Immunol. 2013; 25:152–9.

2). Sahin U, Karikó K, Tüereci Ö. mRNA-based thera- peutics – developing a new class of drugs. Nat Rev Drug Discov. 2014; 13:759–80.
3). Smith ML, Corbo T, Bernales J, Lindbo JA, Pogue GP, Palmer KE, et al. Assembly of trans-encapsidated recombinant viral vectors engineered from Tobacco mosaic virus and Semliki Forest virus and their evaluation as immunogens. Virology. 2007; 358:321–33.
4). Azizgolshani O, Garmann RF, Cadena-Nava R, Knobler CM, Gelbart WM. Reconstituted plant viral capsids can release genes to mammalian cells. Virology. 2013; 441:12–7.
5). Shin HI, Kim IC, Cho TJ. Replication and encapsidation of recombinant Turnip yellow mosaic virus RNA. BMB Rep. 2008; 41:739–44.
6). Dreher TW. Turnip yellow mosaic virus: transfer RNA mimicry, chloroplasts and a C-rich genome. Mol Plant Pathol. 2004; 5:367–75.
7). Kim HB, Kim DY, Cho TJ. Replication and packaging of Turnip yellow mosaic virus RNA containing Flock house virus RNA1 sequence. BMB Rep. 2014; 47:330–5.
8). Ball LA. Requirements for the self-directed replication of Flock house virus RNA1. J Virol. 1995; 69:720–7.
9). Price BD, Ruekert RR, Ahlquist P. Complete replication of an animal virus and maintenance of expression vectors derived from it in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A. 1996; 93:9465–70.
10). Selling BH, Allison RF, Kaesberg P. Genomic RNA of an insect virus directs synthesis of infectious virions in plants. Proc Natl Acad Sci U S A. 1990; 87:434–8.
11). Johnson KL, Ball LA. Induction and maintenance of autonomous Flock house virus RNA1 replication. J Virol. 1999; 73:7933–42.
12). Li H, Li WX, Ding SW. Induction and suppression of RNA silencing by an animal virus. Science. 2002; 296:1319–21.
13). Shi BJ, Ding SW, Symons RH. Plasmid vector for cloning infectious cDNAs from plant RNA viruses: high infectivity of cDNA clones of tomato aspermy cucumovirus. J Gen Virol. 1997; 78:1181–5.
14). Xiang C, Han P, Lutziger I, Wang K, Oliver DJ. A mini binary vector series for plant transformation. Plant Mol Biol. 1999; 40:711–7.
15). Cho TJ, Dreher TW. Encapsidation of genomic but not subgenomic Turnip yellow mosaic virus RNA by coat protein provided in trans. Virology. 2006; 356:126–35.
16). Perrotta AT, Shih I, Been MD. Imidazole rescue of a cytosine mutation in a self-cleaving ribozyme. Science. 1999; 286:123–6.
17). Albariño CG, Price BD, Eckerle LD, Ball LA. Characterization and template properties of RNA dimers generated during Flock house virus RNA replication. Virology. 2001; 289:269–82.
18). Annamalai P, Rofail F, DeMason DA, Rao AL. Replication-coupled packaging mechanism in positive-strand RNA viruses: Synchronized coexpression of functional multigenome RNA components of an animal and a plant virus in Nicotiana benthamiana cells by agroinfiltraion. J Virol. 2008; 82:1484–95.
19). Ball LA, Li Y. Cis-Acting requirements for the replication of Flock house virus RNA2. J Virol. 1993; 67:3544–51.
20). Eckerle LD, Albariño CG, Ball LA. Flock house virus subgenomic RNA3 is replicated and its replication correlates with transactivation of RNA2. Virology. 2003; 317:95–108.
Go to :
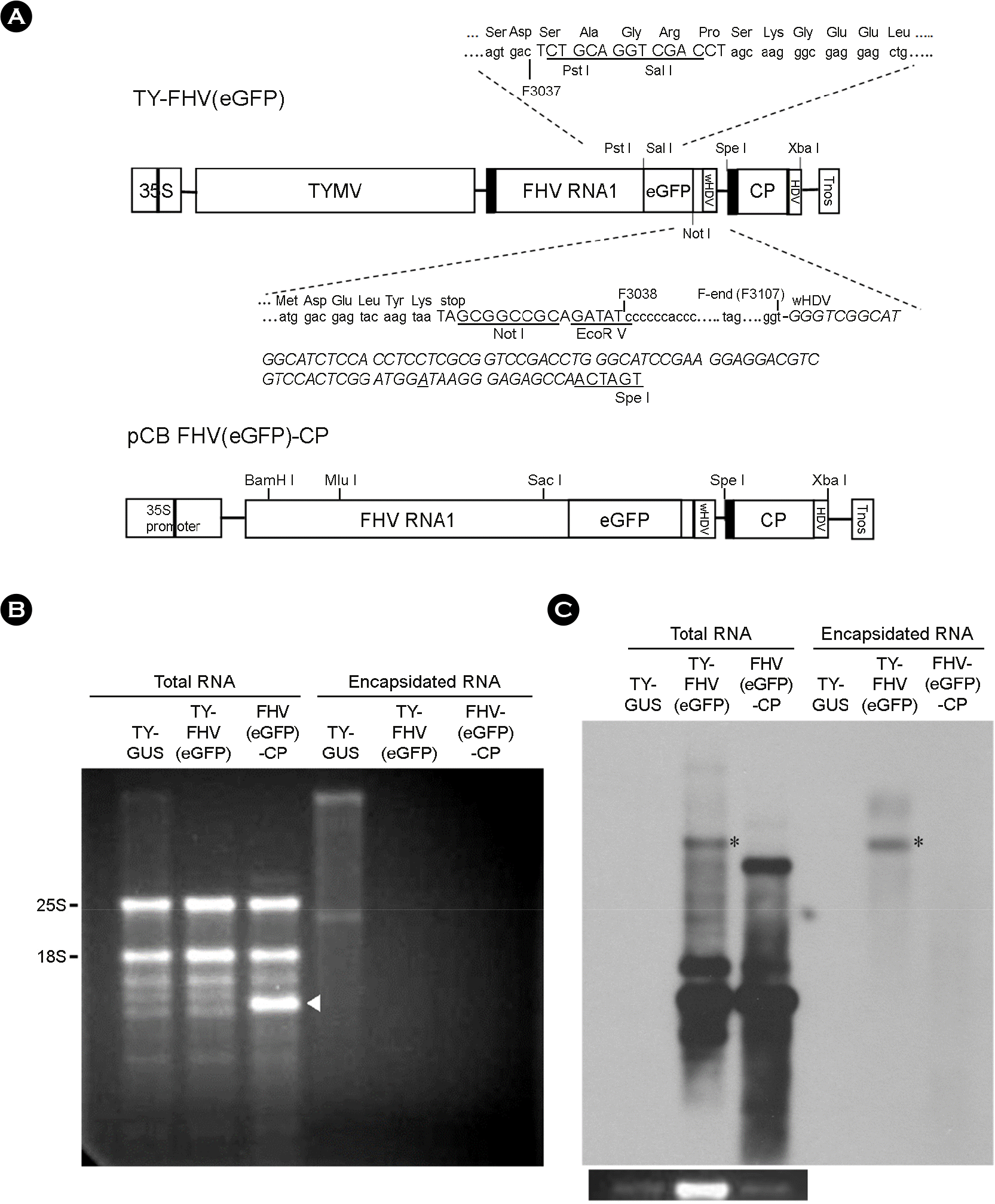 | Figure 1.Replication and encapsidation of recombinant FHV RNA1. (A) DNA constructs. Preparation of TY-FHV(eGFP) was described previously (7). Briefly, FHV RNA1 sequence was placed downstream of the tymobox (black box), a sgRNA promoter. eGFP sequence was inserted into the B2 ORF of the FHV genome. The ribozyme sequence derived from the HDV is italicized. The nucleotide A that makes the ribozyme weak is underlined. A portion of the TY-FHV(eGFP), the DNA comprising the FHV RNA1 sequence, eGFP gene, HDV ribozyme, and TYMV CP sequence, was placed downstream of double 35S promoters of CaMV in pCB 302-3, yielding the pCB FHV(eGFP)-CP construct (for details, see MATERIALS AND METHODS). (B) Agarose gel electrophoresis. Seven days after agroinfiltration of N. benthamiana leaves with various FHV constructs, total RNA was extracted. Encapsidated RNA samples were prepared by incubating leaf extracts with RNase A, followed by phenol/chloroform extraction and ethanol precipitation. RNA samples were size-fractionated in a 1% agarose gel and stained with EtBr. 25S and 18S rRNAs are indicated. (C) Northern blot analysis of FHV RNA replication. 500 ng of total RNA and equivalent amounts of encapsidated RNA were size-fractionated in a 1% agarose gel and examined by northern blot analysis, using the DIG-labeled probe targeting FHV sgRNA. The blots were developed by chemiluminescent immunodetection of DIG. The panel below the northern blot shows 25S rRNA stained with EtBr. Agroinfiltration and northern blot analysis were carried out in triplicate, and a representative blot is shown. TY-GUS construct containing a β-glucuronidase gene (5) was included in this experiment as a negative control, since the FHV probe does not hybridize to the TY-GUS RNA. |
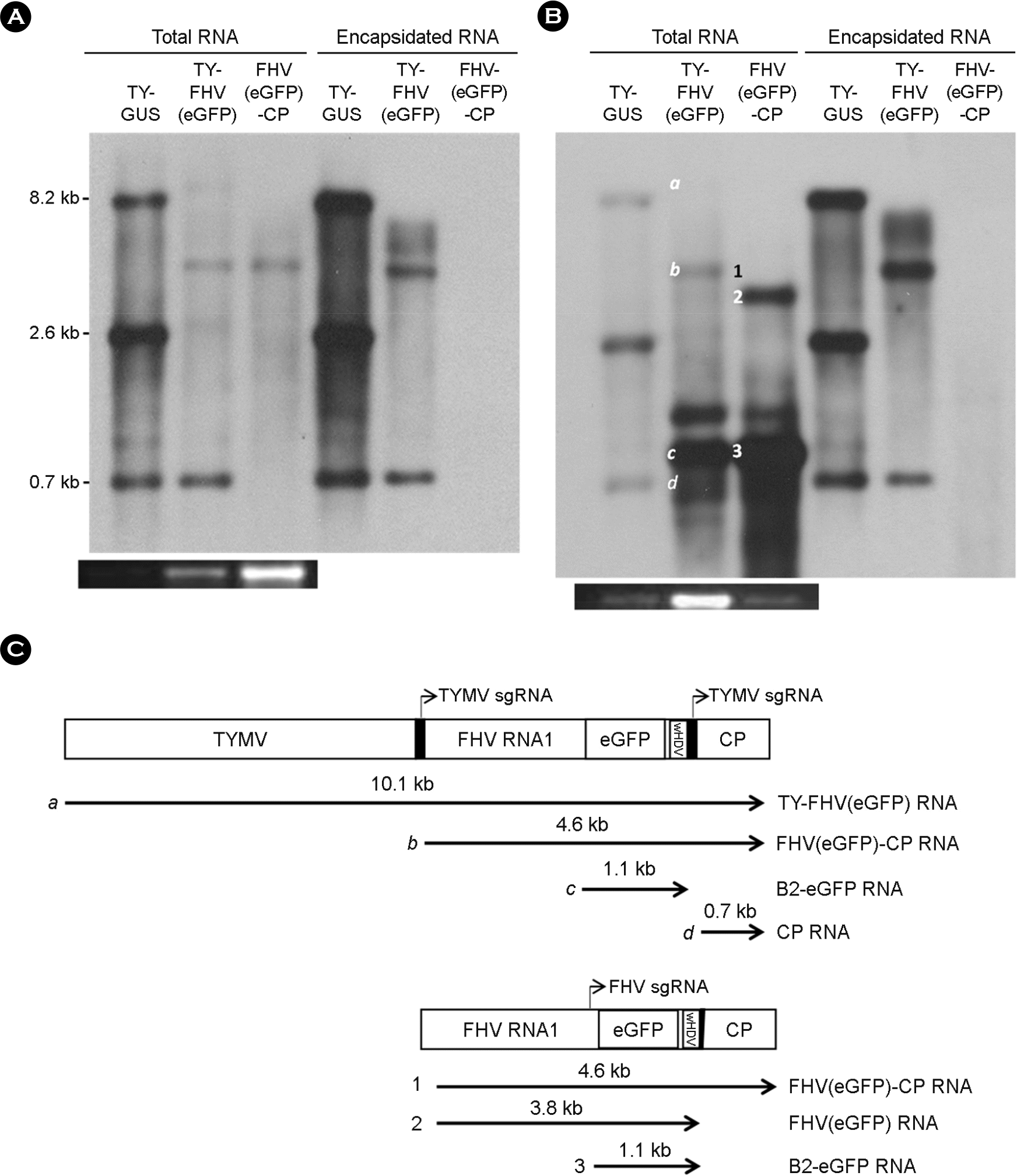 | Figure 2.Analysis of replication products. Northern blot analysis of FHV RNA replication (A & B) was done as described in Figure 1C, using the DIG-labeled probe targeting the TYMV CP sequence (A) or a mixture of probes targeting the FHV sgRNA and TYMV CP probes (B). Experiments were performed in triplicate, yielding similar results. C) Schematic representation of the FHV RNA replication products. The sgRNA promoter tymobox is represented as a black box. Four RNA species (a, b, c, and d) from TY-FHV (eGFP) and three species (1, 2, and 3) from pCB FHV (eGFP)-CP, which were identified by the size and the results of hybridization, are depicted below the blots. TY-GUS RNA was included as a size marker. |
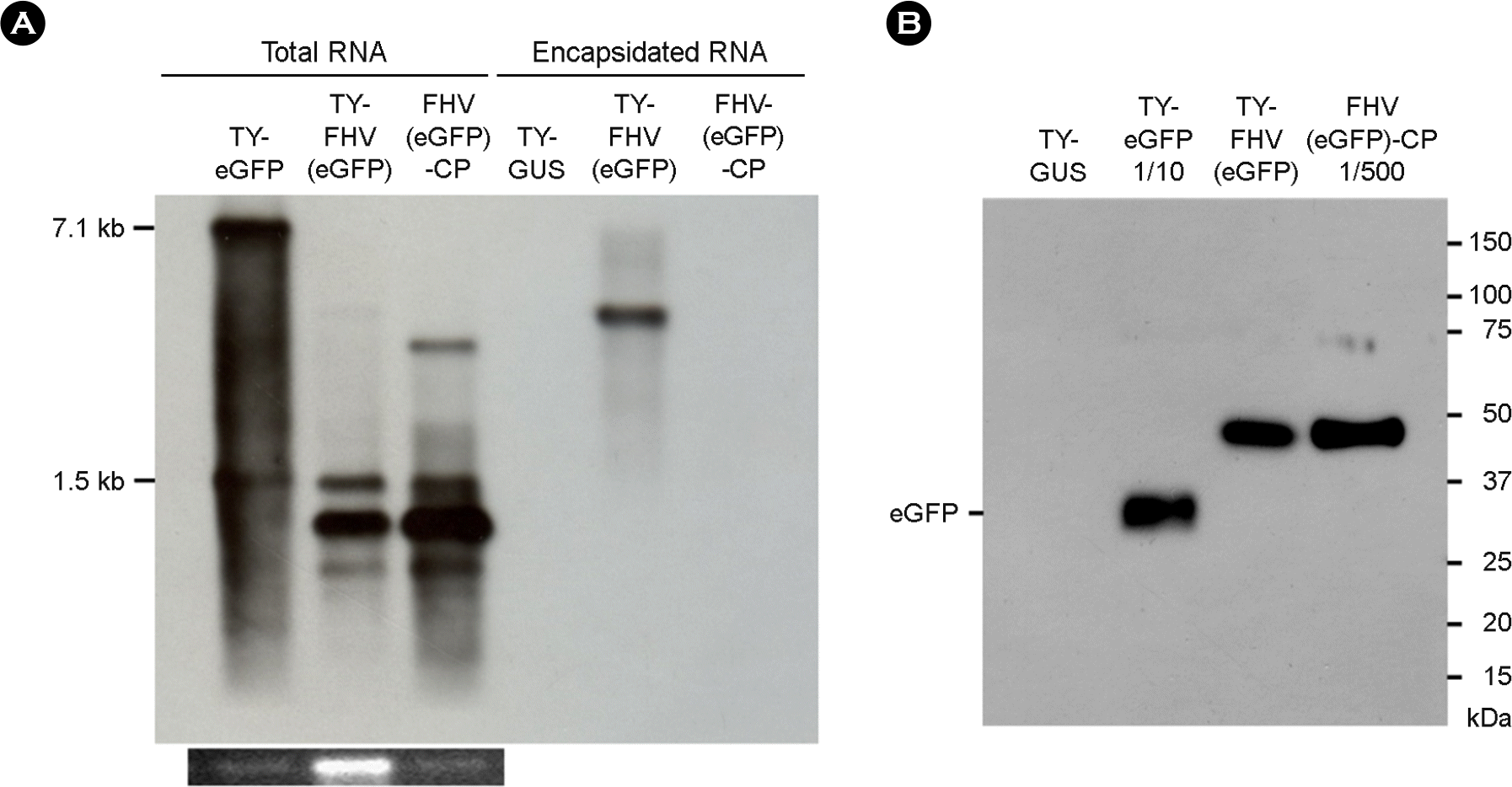 | Figure 3.Analysis of eGFP expression. (A) Northern blot analysis of eGFP expression. Total or encapsidated RNAs were examined as described in Figure 1, using the DIG-labeled probe targeting the eGFP gene. (B) Western blot analysis of eGFP expression. 10 μl of leaf extracts was electrophoresed in a 12.5% SDS-polyacrylamide gel, and the proteins were transferred to nitrocellulose membrane. For TY-eGFP and FHV (eGFP)-CP, diluted samples (1:10 and 1:500 dilution, respectively) were used. eGFP was detected using an anti-eGFP antibody and anti-rabbit HRP conjugate. Experiments were performed in triplicate. |
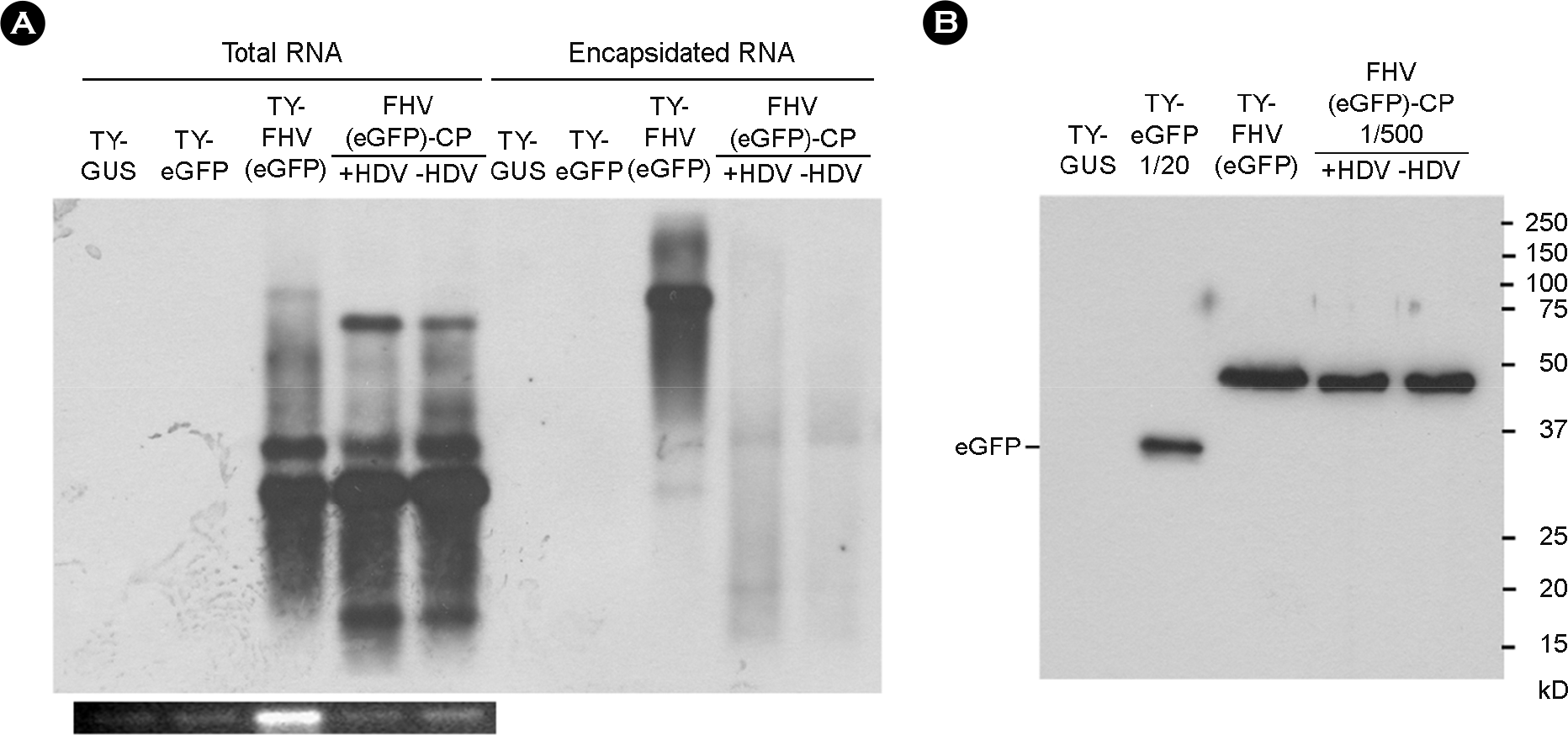 | Figure 4.Replication of recombinant FHV RNA1 lacking HDV ribozyme sequence. (A) Northern blot analysis of FHV RNA1 replication. Total or encapsidated RNAs were examined as described in Fig. 1, using the DIG-labeled probe targeting the FHV sgRNA. (B) Western blot analysis of eGFP expression. 10 μl of leaf extracts was electrophoresed in a 12.5% SDS-polyacrylamide gel, and the proteins were transferred to nitrocellulose membrane. For TY-eGFP, the protein sample was diluted to 1:20. For FHV(eGFP)-CP and FHV(eGFP)ΔHDV-CP, the samples were diluted to 1:500. eGFP was detected as described in Figure 3B. Experiments were performed in triplicate. |
 XML Download
XML Download