

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Turnip yellow mosaic virus (TYMV) is a spherical plant virus that has a single 6.3 kb positive strand RNA as a genome. Previously, we have made the recombinant TYMV construct containing a 0.7 kb eGFP gene or a 1.8 kb GUS gene. The genomic RNAs from these constructs were efficiently encapsidated. To examine in more detail whether size constraint exists for replication and packaging of TYMV, we have inserted into the TY-GUS an extra sequence derived from either eGFP or GUS. We also made a recombinant containing RNA1 sequence of Flock house virus. These TYMV recombinants were introduced into Nicotiana benthamiana leaves by agroinfiltration. Northern blot analysis of the viral RNAs in the agroinfiltrated leaves showed that the genomic RNA band from the recombinant TYMV became weaker as longer sequence was inserted. The result also showed that the efficiency of genomic RNA encapsidation decreased sharply when an extra sequence of 2.2 kb or more was inserted. In contrast, the recombinant subgenomic RNA containing an extra sequence of up to 3.2 kb was efficiently encapsidated. Overall, these results show that size constraint exists for replication and encapsidation of TYMV RNA.
Go to : 
REFERENCES
1). Dreher TW. Turnip yellow mosaic virus: transfer RNA mimicry, chloroplasts and a C-rich genome. Mol Plant Pathol. 2004; 5:367–75.

2). Canady MA, Larson SB, Day J, McPherson A. Crystal structure of turnip yellow mosaic virus. Nat Struct Biol. 1996; 3:771–81.
3). Mellema JR, Benicourt C, Haenni AL, Noort A, Pleij CW, Bosch L. Translational studies with turnip yellow mosaic virus RNAs isolated from major and minor virus particles. Virology. 1979; 96:38–46.
4). Rao ALN. Genome packaging by spherical plant RNA viruses. Annu Rev Phytopathol. 2006; 44:61–87.
5). Qu F, Morris TJ. Encapsidation of turnip crinkle virus is defined by a specific packaging signal and RNA size. J Virol. 1997; 71:1428–35.
6). Shin HI, Kim IC, Cho TJ. Replication and encapsidation of recombinant Turnip yellow mosaic virus RNA. BMB Rep. 2008; 41:739–44.
7). Cho TJ, Dreher TW. Encapsidation of genomic but not subgenomic turnip yellow mosaic virus RNAs by coat protein provided in trans. Virology. 2006; 356:126–35.
8). Voinnet O, Rivas S, Mestre P, Baulcombe D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003; 33:949–56.
9). Albarino CG, Price BD, Eckerle LD, Ball LA. Characterization and template properties of RNA dimers generated during Flock house virus RNA replication. Virology. 2001; 289:269–82.
10). Matthews REF. Tymovirus group. Plant Virology. 3rd ed.Academic Press;1991. p. p231–39.
11). Hellendoorn K, Michiels PJ, Buitenhuis R, Pleij CW. Protonatable hairpins are conserved in the 5′-untranslated region of tymovirus RNAs. Nucl Acids Res. 1996; 24:4910–7.
12). Hellendoorn K, Verlaan PWG, Pleij CWA. A functional role for the conserved protonatable hairpins in the 5′ untranslated region of turnip yellow mosaic virus RNA. J Virol. 1997; 71:8774–9.
13). Bink HHJ, Hellendoorn K, van der Meulen J, Pleij CWA. Protonation of non-Watson-Crick base pairs and encapsidation of turnip yellow mosaic virus RNA. Proc Natl Acad Sci. 2002; 99:13465–70.
14). Shin HI, Tzanetakis IE, Dreher TW, Cho TJ. The 5′-UTR of Turnip yellow mosaic virus does not include a critical encapsidation signal. Virology. 2009; 387:427–35.
15). Choi YG, Dreher TW, Rao ALN. tRNA elements mediate the assembly of an icosahedral RNA virus. Proc Natl Acad Sci. 2002; 99:655–60.
16). Shin HI, Kim HY, Cho TJ. The Pro/Hel region is indispensable for packaging non-replicating Turnip yellow mosaic virus RNA, but not replicating viral RNA. Mol. Cells. 2010; 29:463–9.
17). Shin HI, Cho NJ, Cho TJ. Role of 5′-UTR hairpins of the Turnip yellow mosaic virus RNA in replication and systemic movement. BMB Rep. 2008; 41:778–83.
Go to :
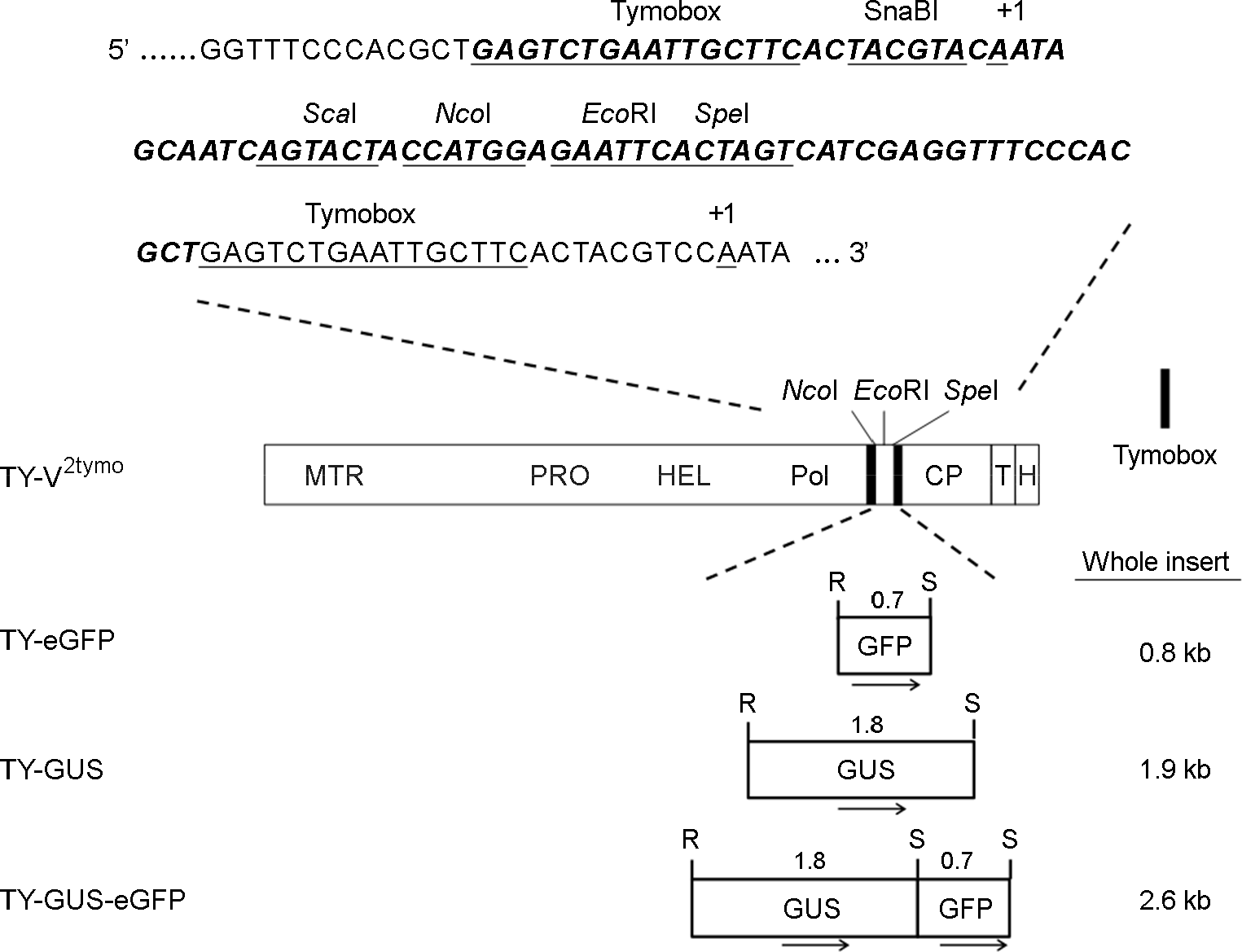 | Figure 1.Recombinant TYMV constructs. TY-V2tymo is an expression vector that has an extra tymobox, a subgenomic RNA promoter, and a multiple cloning site (6). The methyltransferase (MTR), protease (PRO), helicase (HEL), and polymerase (POL) domains of TYMV genome are indicated. T and H represent the tRNA-like structure and hepatitis delta virus ribozyme, respectively. The inserted sequence is shown in bold and italic letters. Transcription start site is indicated by +1. TY-eGFP, TY-GUS, and TY-GUS-eGFP were prepared by inserting the foreign sequence into the EcoRI/SpeI sites of TY-V2tymo. R and S represent EcoRI and SpeI restriction sites, respectively. The sizes of the whole insert are shown on the right. The arrows below the inserted genes represent the direction of transcription. The numeral above the inserted sequence indicates the size of the gene. |
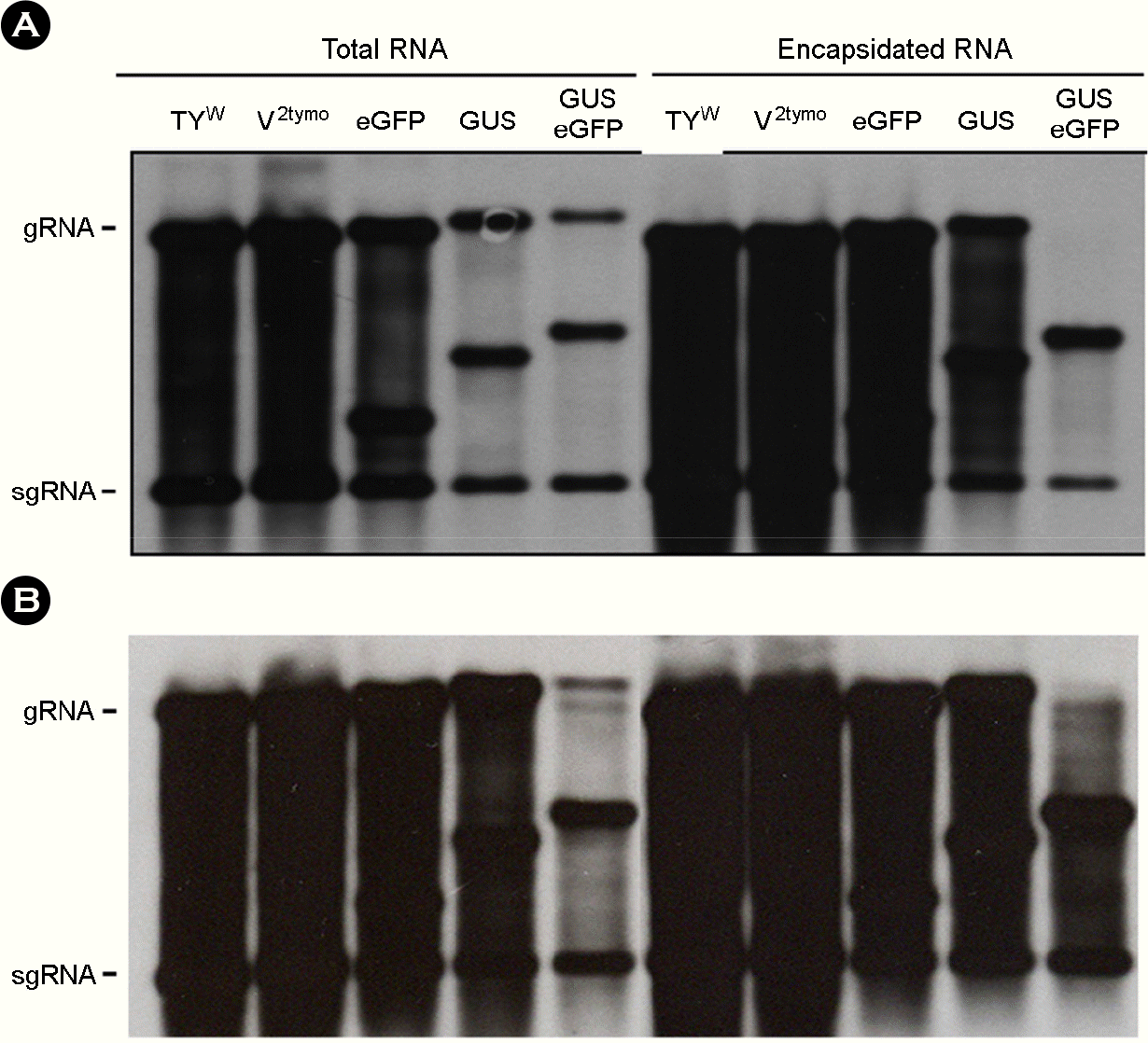 | Figure 2.Replication and packaging of recombinant TYMV constructs. Recombinant TYMV constructs were agroinfiltrated into N. benthamiana plants. A week after agroinfiltration, total RNA was extracted from the agroinfiltrated leaf. For encapsidation assay, leaf extracts were treated with RNase A (5 μg/ml), followed by phenol/chloroform extraction. Experiments were repeated at least two times. RNA samples were size-fractionated by agarose gel electrophoresis and analyzed by Northern blot hybridization using a DIG-labeled CP ORF DNA as a probe. Two representative blots are shown (A and B). The positions of genomic RNA (gRNA) and subgenomic RNA (sgRNA) are indicated. |
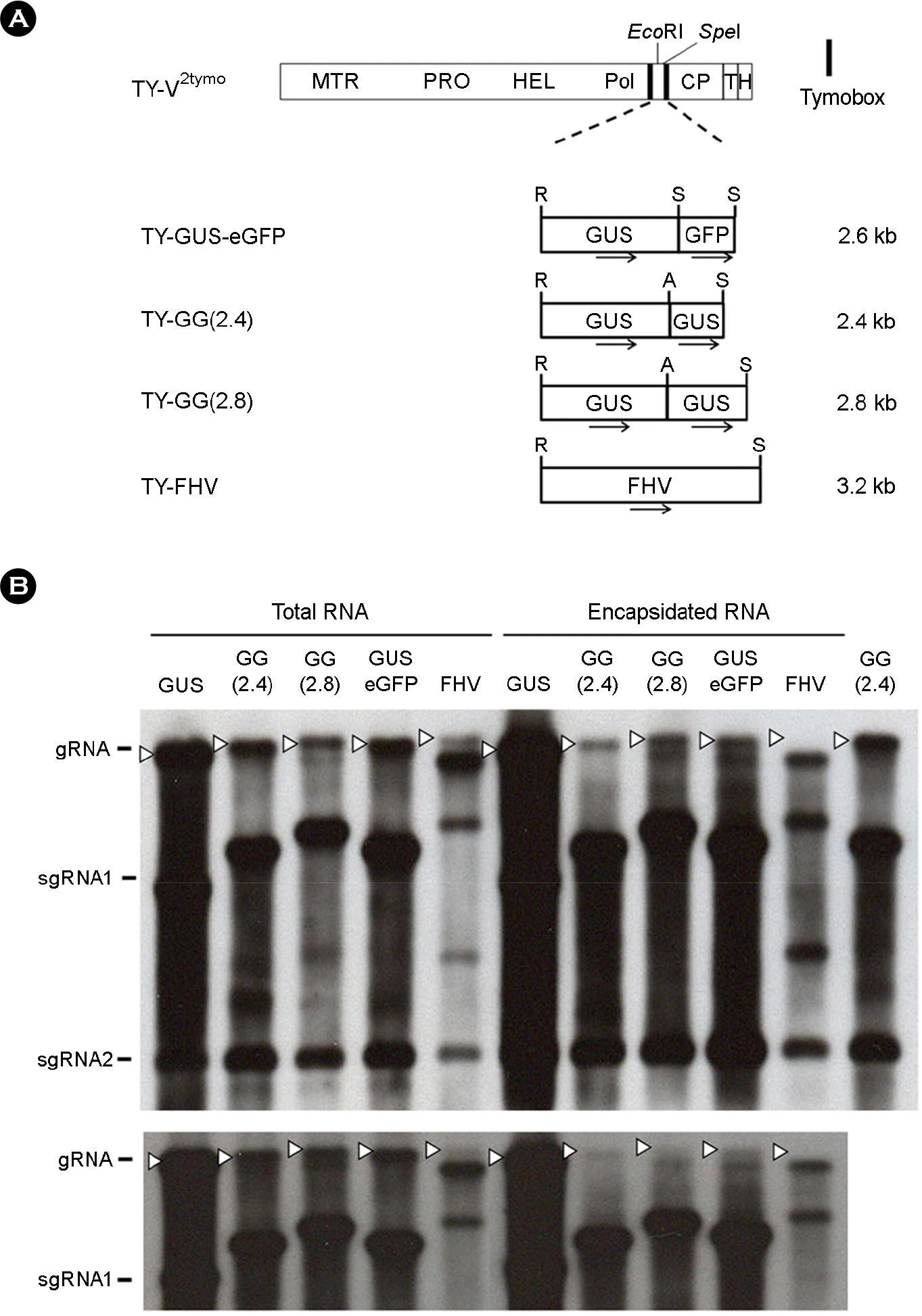 | Figure 3.Replication and packaging of the recombinant TYMV whose insert is equal to or larger than 2.4 kb. (A) TYMV constructs. TY-GG(2.4) and TY-GG(2.8) constructs were prepared by inserting 0.8 kb and 1.2 kb extra GUS sequence, respectively, into the AscI/SpeI sites of the TY-GUS(1.6), where AscI recognition sequence was introduced at the 3′-end of the insert (see MATERIALS AND METHODS for details). TY-FHV construct contains a RNA1 sequence of Flock house virus. The sizes of the inserts are represented on the right of the constructs. Orientation of the inserted gene sequences are indicated by arrows. (B) Northern analysis of the replication and packaging of the recombinant TYMV RNAs. Total RNA and encapsidated RNA samples were analyzed by Northern blot hybridization, as described in Fig. 2. Two representative blots are shown. The bottom panel represents an upper part of the gel showing the gRNAs and the larger sgRNAs. The rightmost sample in the top panel is a total RNA sample from TY-GG(2.4), which was included as a blotting control. gRNAs are indicated by white arrowheads. |
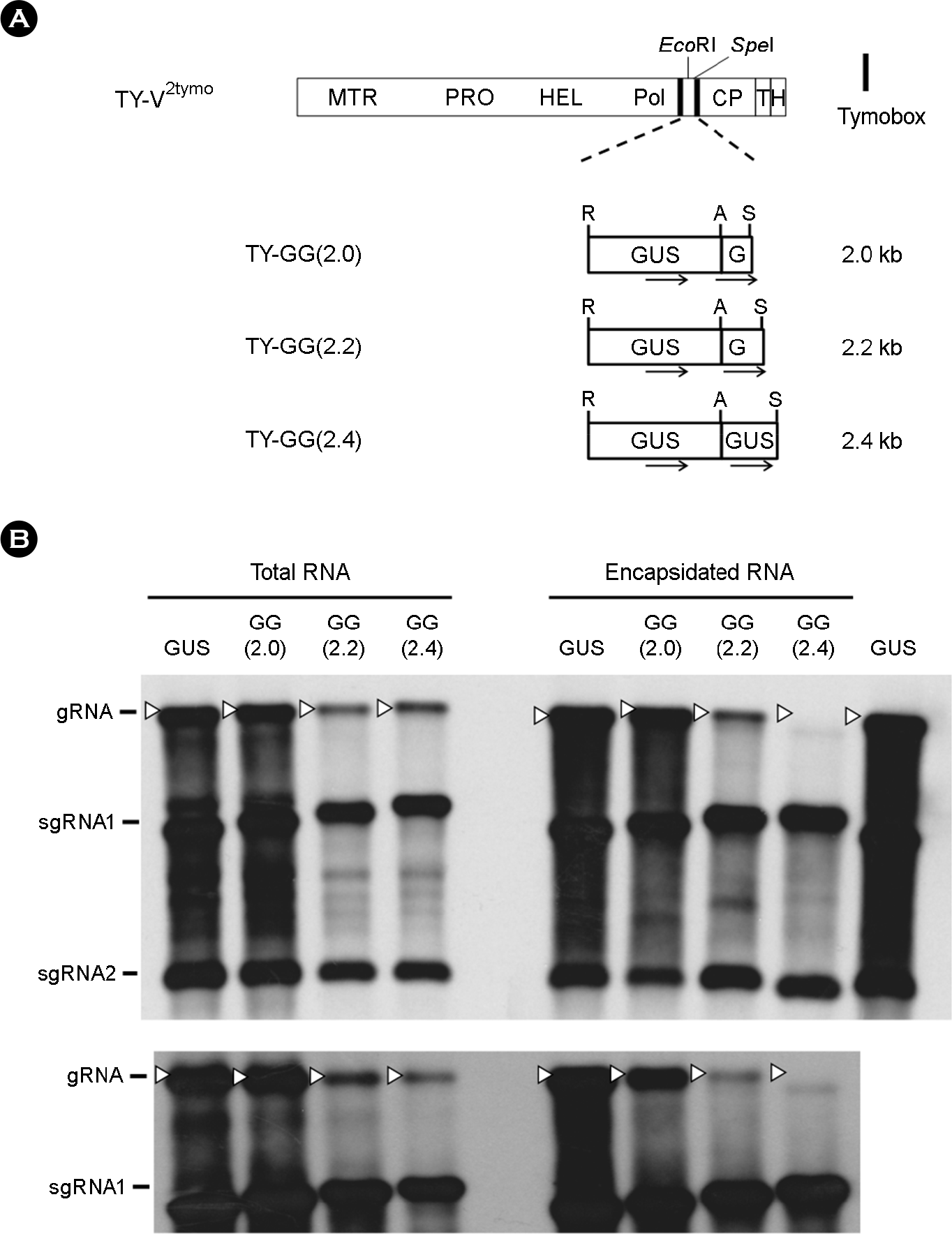 | Figure 4.Replication and packaging of the recombinant TYMV whose insert is smaller than 2.4 kb. (A) TYMV constructs. TY-GG (2.0) and TY-GG(2.2) constructs were prepared by inserting 0.4 kb and 0.6 kb extra GUS sequence, respectively, into the AscI/SpeI sites of the TY-GUS(1.6). (B) Northern analysis of the replication and packaging of the recombinant TYMV RNAs. Total RNA and encapsidated RNA samples were analyzed by Northern blot hybridization, as described in Fig. 2. The rightmost TY-GUS sample in the top panel was included as a blotting control. gRNAs are indicated by white arrowheads. |
 XML Download
XML Download