

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
15-deoxy-delta12,14 prostaglandin J2 (15d-PGJ2) may hold promise in treatment of the pathologies associated with human immunodeficiency virus (HIV) infection of the central nervous system. However, its precise role and neuroprotective mechanism in the hippocampus remain poorly understood. In the present study, rat hippocampal slices were stimulated with HIV-1 Tat protein to investigate the protective role of 15d-PGJ2 on the hippocampal cytotoxicity. Full-length HIV-1 Tat protein (Tat1-86), but neither its Tat32-62 nor Tat30-86 fragment, significantly induced cytotoxicity in the hippocampus, the brain region most commonly damaged in HIV-associated dementia. This Tat-induced cytotoxicity was associated with inactivation of MEK/extracellular signal-regulated kinase (ERK) signaling pathway. In contrast, Tat1-86 did not alter Wnt signaling pathway necessary for cell survival. Pretreatment of slices with 15d-PGJ2 markedly reduced Tat-driven cytotxicity. Interestingly, this reduction was accompanied by suppression of ERK inactivation in response to Tat. Moreover, the inhibition of the MEK/ERK pathway with SL327 enhanced the Tat-induced cytotoxicity, confirming the ERK-dependent mechanism of Tat-driven cytotoxicity. Collectively, these data demonstrate that the protective action of 15d-PGJ2 against the hippocampal cytotoxicity upon Tat stimulation is exerted through suppression of Tat-mediated ERK1/2 inactivation.
Go to : 
REFERENCES
1). Gannon P, Khan MZ, Kolson DL. Current understanding of HIV-associated neurocognitive disorders pathogenesis. Curr Opin Neurol. 2011; 24:275–83.

2). Irish BP, Khan ZK, Jain P, Nonnemacher MR, Pirrone V, Rahman S, et al. Molecular mechanisms of neuro-degenerative diseases induced by human retroviruses: a review. Am J Infect Dis. 2009; 5:231–58.
3). Wright E. Neurocognitive impairment and neuroCART. Curr Opin HIV AIDS. 2011; 6:303–8.
4). Romani B, Engelbrecht S, Glashoff RH. Functions of Tat: the versatile protein of human immunodeficiency virus type 1. J Gen Virol. 2010; 91:1–12.
5). Kaul M, Lipton SA. Mechanisms of neuronal injury and death in HIV-1 associated dementia. Curr HIV Res. 2006; 4:307–18.
6). Khiati A, Chaloin O, Muller S, Tardieu M, Horellou P. Induction of monocyte chemoattractant protein-1 (MCP-1/CCL2) gene expression by human immunodeficiency virus-1 Tat in human astrocytes is CDK9 dependent. J Neurovirol. 2010; 16:150–67.
7). Turchan-Cholewo J, Dimayuga VM, Gupta S, Gorospe RM, Keller JN, Bruce-Keller AJ. NADPH oxidase drives cytokine and neurotoxin release from microglia and macrophages in response to HIV-Tat. Antioxid Redox Signal. 2009; 11:193–204.
8). Williams R, Yao H, Peng F, Yang Y, Bethel-Brown C, Buch S. Cooperative induction of CXCL10 involves NADPH oxidase: implications for HIV dementia. Glia. 2010; 58:611–21.
9). Del Valle L, Croul S, Morgello S, Amini S, Rappaport J, Khalili K. Detection of HIV-1 Tat and JCV capsid protein, VP1, in AIDS brain with progressive multifocal leukoencephalopathy. J Neurovirol. 2000; 6:221–8.
10). Yoon CH, Woo JY, Bae YS. HIV-1 Infection causes intracellular expression of p53, which induces PKR expression, followed by inhibition of HIV-1 Tat activity. J Bacteriol Virol. 2004; 34:157–66.
11). Kalantari P, Narayan V, Henderson AJ, Prabhu KS. 15-Deoxy-Delta12,14-prostaglandin J2 inhibits HIV-1 transactivating protein, Tat, through covalent modification. FASEB J. 2009; 23:2366–73.
12). Glass CK, Saijo K, Winner B, Marchetto MC, Gage FH. Mechanisms underlying inflammation in neurodegeneration. Cell. 2010; 140:918–34.
13). Lee YJ, Han SB, Nam SY, Oh KW, Hong JT. Inflammation and Alzheimer's disease. Arch Pharm Res. 2010; 33:1539–56.
14). Chong YH, Shin YJ, Lee EO, Kayed R, Glabe CG, Tenner AJ. ERK1/2 activation mediates Abeta oligomer-induced neurotoxicity via caspase-3 activation and tau cleavage in rat organotypic hippocampal slice cultures. J Biol Chem. 2006; 281:20315–25.
15). Lee EO, Kim SE, Park HK, Kang JL, Chong YH. Extracellular HIV-1 Tat upregulates TNF-α dependent MCP-1/CCL2 production via activation of ERK1/2 pathway in rat hippocampal slice cultures: Inhibition by resveratrol, a polyphenolic phytostilbene. Exp Neurol. 2011; 229:399–408.
16). Holopainen IE. Organotypic hippocampal slice cultures: a model system to study basic cellular and molecular mechanisms of neuronal cell death, neuroprotection, and synaptic plasticity. Neurochem Res. 2005; 30:1521–8.
17). Manzoor Z, Koh YS. Mitogen-activated protein kinases in inflammation. J Bacteriol Virol. 2012; 42:189–95.
18). Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science. 2004; 303:1483–7.
19). Cadigan KM, Liu YI. Wnt signaling: complexity at the surface. J Cell Sci. 2006; 119:395–402.
20). Sharma A, Hu XT, Napier TC, Al-Harthi L. Methamphetamine and HIV-1 Tat down regulate β-catenin signaling: implications for methampetamine abuse and HIV-1 co-morbidity. J Neuroimmune Pharmacol. 2011; 6:597–607.
21). Aksenov MY, Aksenova MV, Nath A, Ray PD, Mactutus CF, Booze RM. Cocaine-mediated enhancement of Tat toxicity in rat hippocampal cell cultures: the role of oxidative stress and D1 dopamine receptor. Neurotoxicology. 2006; 27:217–28.
22). Aksenova MV, Aksenov MY, Adams SM, Mactutus CF, Booze RM. Neuronal survival and resistance to HIV-1 Tat toxicity in the primary culture of rat fetal neurons. Exp Neurol. 2009; 215:253–63.
23). Kruman II, Nath A, Mattson MP. HIV-1 protein Tat induces apoptosis of hippocampal neurons by a mechanism involving caspase activation, calcium overload, and oxidative stress. Exp Neurol. 1998; 154:276–88.
24). Brailoiu GC, Brailoiu E, Chang JK, Dun NJ. Excitatory effects of human immunodeficiency virus 1 Tat on cultured rat cerebral cortical neurons. Neuroscience. 2008; 151:701–10.
25). Nath A, Psooy K, Martin C, Knudsen B, Magnuson DS, Haughey N, et al. Identification of a human immunodeficiency virus type 1 Tat epitope that is neuroexcitatory and neurotoxic. J Virol. 1996; 70:1475–80.
26). Brana C, Biggs TE, Mann DA, Sundstrom LE. A macrophage hippocampal slice co-culture system: application to the study of HIV-induced brain damage. J Neurosci Methods. 1999; 90:7–11.
27). Yao H, Peng F, Dhillon N, Callen S, Bokhari S, Stehno-Bittel L, et al. Involvement of TRPC channels in CCL2-mediated neuroprotection against tat toxicity. J Neurosci. 2009; 29:1657–69.
28). Yao H, Peng F, Fan Y, Zhu X, Hu G, Buch SJ. TRPC channel-mediated neuroprotection by PDGF involves Pyk2/ERK/CREB pathway. Cell Death Differ. 2009; 16:1681–93.
29). Zhu X, Yao H, Peng F, Callen S, Buch S. PDGF-mediated protection of SH-SY5Y cells against Tat toxin involves regulation of extracellular glutamate and intracellular calcium. Toxicol Appl Pharmacol. 2009; 240:286–91.
30). Shi J, Qin X, Zhao L, Wang G, Liu C. Human immunodeficiency virus type 1 Tat induces B7-H1 expression via ERK/MAPK signaling pathway. Cell Immunol. 2011; 271:280–5.
31). Appel S, Mirakaj V, Bringmann A, Weck MM, Grünebach F, Brossart P. PPAR-gamma agonists inhibit toll-like receptor-mediated activation of dendritic cells via the MAP kinase and NF-kappaB pathways. Blood. 2005; 106:3888–94.
32). Chana RS, Lewington AJ, Brunskill NJ. Differential effects of peroxisome proliferator activated receptor-gamma (PPAR gamma) ligands in proximal tubular cells: thiazolidinediones are partial PPAR gamma agonists. Kidney Int. 2004; 65:2081–90.
33). Alvarez-Maqueda M, El Bekay R, Alba G, Monteseirín J, Chacón P, Vega A, et al. 15-Deoxy-Δ12,14-prostaglandin J2 induces heme oxygenase-1 gene expression in a reactive oxygen species-dependent manner in human lymphocytes. J Biol Chem. 2004; 279:21929–37.
34). Kim DH, Kim JH, Kim EH, Na HK, Cha YN, Chung JH, et al. 15-Deoxy-Delta12,14-prostaglandin J2 upregulates the expression of heme oxygenase-1 and subsequently matrix metalloproteinase-1 in human breast cancer cells: possible roles of iron and ROS. Carcinogenesis. 2009; 30:645–54.
35). Chiba T, Ueki S, Ito W, Kato H, Takeda M, Kayaba H, et al. 15-Deoxy-Δ(12,14)-prostaglandin J2 induces IL-8 and GM-CSF in a human airway epithelial cell line (NCI-H292). Int Arch Allergy Immunol. 2009; 149:77–82.
36). Wilmer WA, Dixon C, Lu L, Hilbelink T, Rovin BH. A cyclopentenone prostaglandin activates mesangial MAP kinase independently of PPARγ. Biochem Biophys Res Commun. 2001; 281:57–62.
37). Qin S, McLaughlin AP, De Vries GW. Protection of RPE cells from oxidative injury by 15-deoxydelta12,14-prostaglandin J2 by augmenting GSH and activating MAPK. Invest Ophthalmol Vis Sci. 2006; 47:5098–105.
38). Kim EH, Na HK, Surh YJ. Upregulation of VEGF by 15-deoxy-Delta12,14-prostaglandin J2 via heme oxygenase-1 and ERK1/2 signaling in MCF-7 cells. Ann N Y Acad Sci. 2006; 1090:375–84.
Go to :
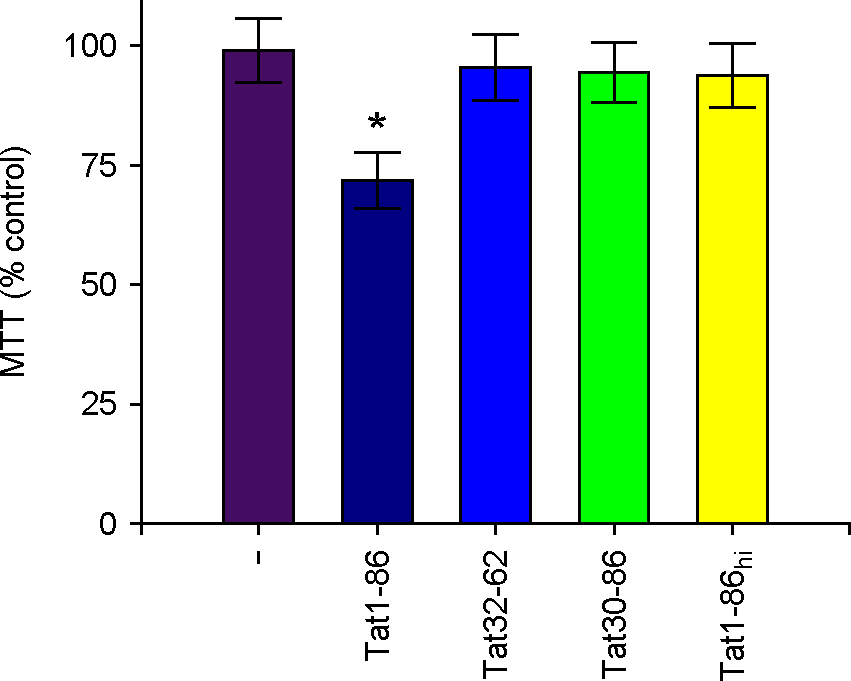 | Figure 1.Influences of Tat on cell toxicity of hippocampal slices. Hippocampal slices were exposed to 1 μM of Tat1-86, Tat fragments, or heat inactivated Tat1-86 (Tat1-86hi) as indicated for 3 days. MTT reduction activity in Tat-treated slices were assayed as described in materials and methods. Values are expressed as the means ± SEM of triplicate experiments. *, p < 0.05 versus vehicle-treated samples. |
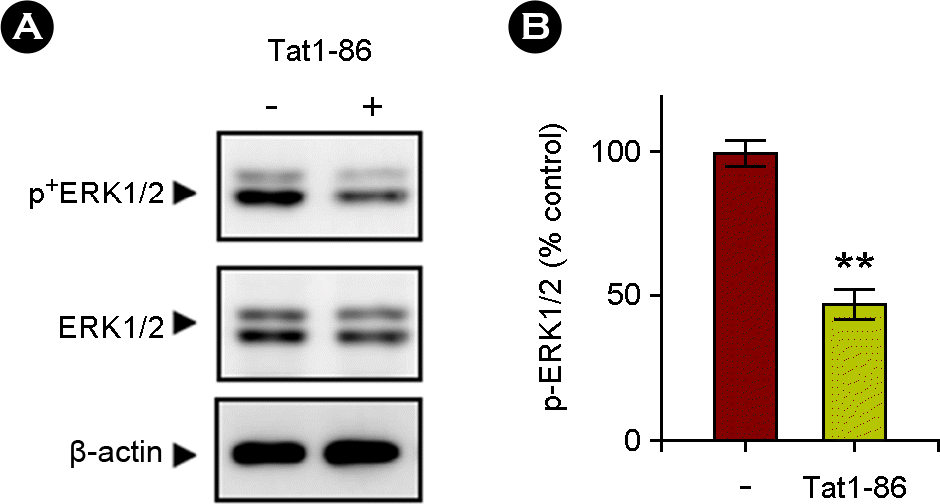 | Figure 2.Tat1-86 decreased the activation of ERK1/2 signaling pathway. The slices were treated for 3 days with Tat1-86 (1 μM) or vehicle only. (A) Immunoblots showing the Tat-induced inactivation of the ERK1/2 signaling pathway in hippocampal slices. Total lysates were analyzed via immunoblotting with phospho-antibody against the activated form of ERK1/2 or anti-total ERK1/2 evidencing decreased ERK1/2 phosphorylation. The blots were stripped and developed with anti-β-actin for equal protein loading. The data represent three independent experiments. (B) A normalized densitometric quantification of ERK1/2 activity against-β-actin for equal protein loading. The results of triplicate experiments are expressed as the means ± SEM. **, p < 0.01 versus vehicle-treated samples. |
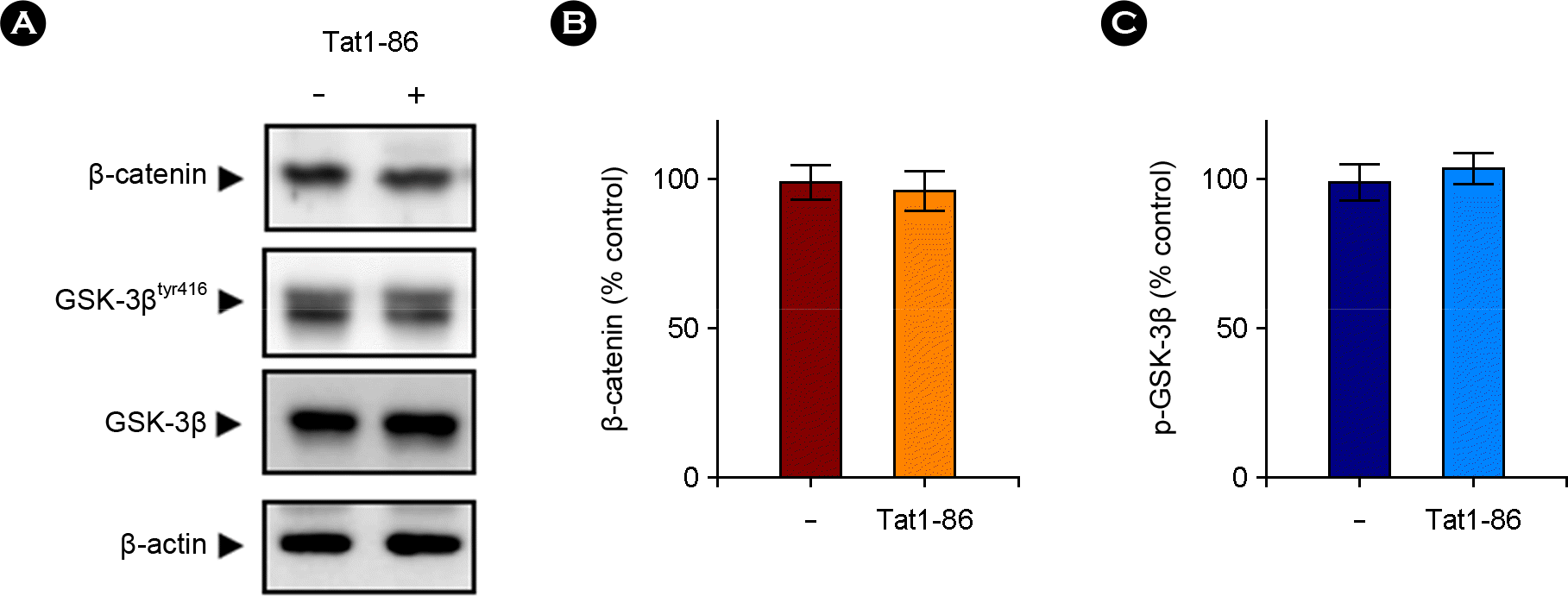 | Figure 3.Effects of Tat1-86 on the Wnt signaling pathway in the hippocampus. (A) Western blotting was used to measure the proteolysis of β-catenin and GSK-3β using total lysates of hippocampal slices treated for 3 days with Tat1-86 (1 μM) or vehicle only as described for Fig. 2. Representative western blots showed that β-catenin level was not changed after Tat treatment for 3 days. Similarly, levels of the active form of GSK-3β (phosphorylated GSK-3βtyr216) and non-phosphorylated GSK-3β were not altered. (B and C) Quantitative analysis of A, showing the levels of β-catenin (B) and GSK-3β (C) normalized to β-actin. Numbers represent the means ± SEM (n = 3). Representative gels from three experiments are shown. |
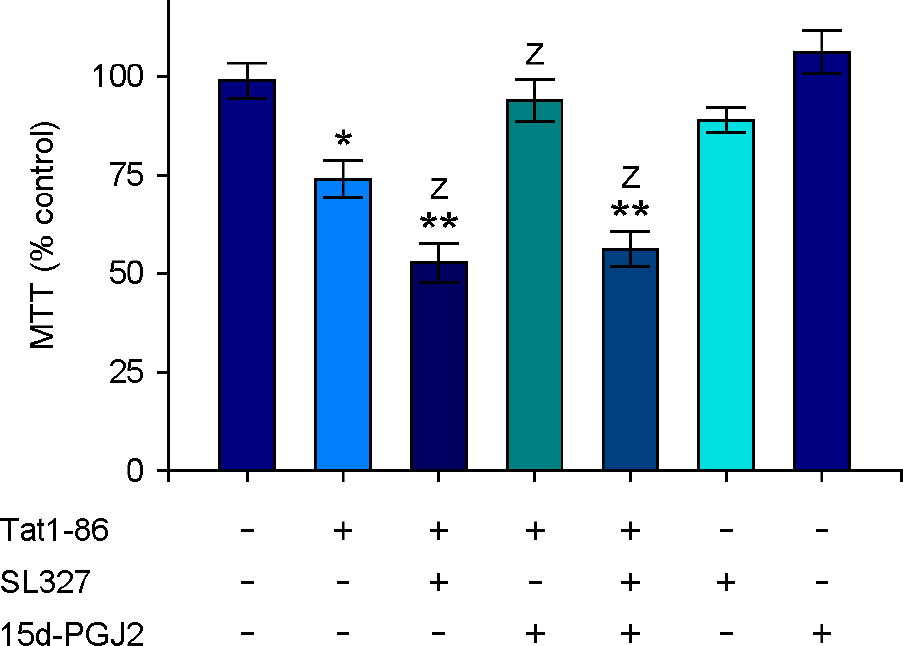 | Figure 4.15d-PGJ2 attenuated Tat-dependent cytotoxicity in the hippocampus. Hippocampal slices were pretreated with 15d-PGJ2 (5 μM) and/or SL327 (10 μM) for 1 h and then stimulated with Tat1-86 (1 μM) for 3 days. MTT reduction activity in Tat-treated slices was measured as described in Fig. 1. Values are expressed as the means ± SEM (n=5). *, p < 0.05; **, p < 0.01 versus vehicle-treated samples; Z, p < 0.05 versus Tat-treated samples. |
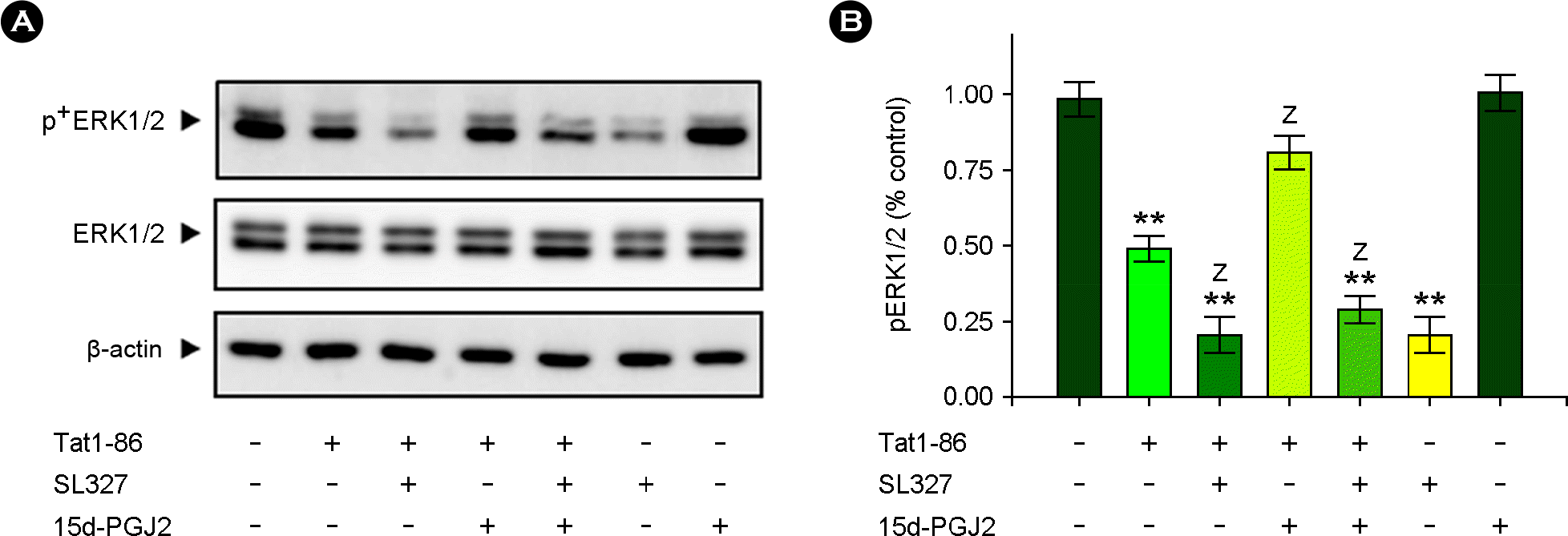 | Figure 5.15d-PGJ2 exerted its protective action against Tat-induced cytotoxicity through ERK1/2 activation. Hippocampal slices were pretreated with 15d-PGJ2 (5 μM) and/or SL327 (10 μM) for 1 h and then stimulated with Tat1-86 (1 μM) for 3 days. Total lysates were analyzed via immunoblotting as described in Fig. 2. (A) Western blots showed that 15d-PGJ2 suppressed Tat-induced inactivation of the ERK1/2 pathway in hippocampal slices. (B) A normalized densitometric quantification of ERK1/2 activity against β-actin for equal protein loading. The results of experiments are expressed as the means ± SEM (n= 5). **, p < 0.01 versus vehicle-treated samples; Z, p < 0.05 versus Tat-treated samples. |
 XML Download
XML Download