

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Epstein Barr Virus (EBV), a member of the human γ-herpesvirus family, is one of the most common viruses in human and is causally associated with lymphoid and epithelial malignancies such as Burkitt's lymphoma (BL), T-cell lymphoma, Hodgkin's Disease, and Nasopharyngeal Carcinoma (1, 2). Latent-infection membrane protein 1 (LMP1), expressed in most EBV-associated malignancies, is an EBV oncogenic protein that is essential for EBV-mediated primary B lymphocyte transformation into proliferating lymphoblastoid cell lines (LCLs) (1, 2). LMP1 is composed of a 24-amino acids (aa) cytosolic N-terminus, 162-aa six hydrophobic transmembrane domains and a 200-aa cytosolic C-Terminus (reviewed in (3)). Using the transmembrane domains, LMP1 aggregates in plasma membrane lipid rafts and constitutively activates NF-κB, p38 mitogen-activated protein kinase (MAPK), c-Jun N-terminal kinase (JNK), and interferon regulatory factor 7 (IRF7) through two C-terminal cytoplasm signaling domains referred to as C-Terminal Activation Regions (CTAR) 1 and 2 (4~11). LMP1-mediated NF-κB activation is essential for EBV-transformed LCL survival (12, 13).
NF-κB is a transcription factor family regulating expression of the genes involved in cell proliferation, differentiation, and survival (reviewed in (14, 15)). The NF-κB family consists of p65/RelA, RelB, c-Rel, p105/p50 and p100/p52 and forms homo-or hetero-dimers to transactivate various genes (14~16). There are two types of NF-κB activation pathways. One is a canonical pathway for NF-κB activation that involves the p65/p50 complexes that are retained in the cytoplasm by inhibitor of κB (IκB) proteins. In response to various stimuli, IκB proteins are phosphorylated by IκB kinase β (IKKβ) and degraded by the ubiquitin-proteasome pathway allowing the p65/p50 complexes to translocate into the nucleus. The other is an alternative pathway for NF-κB activation that involves NF-κB inducing kinase (NIK)-and IKKα-mediated proteolytic processing of p100 into p52 and translocation of the RelB/p52 complexes into the nucleus (14, 15). LMP1 can activate both the canonical and the alternative pathways for NF-κB activation (14, 15). LMP1 CTAR1 engages TRAF1, 2, 3, and 5 through a consensus PXQXT motif found in the CD40 and activates the alternative pathway for NF-κB activation. CTAR2 engages TNFR-associated death domain protein (TRADD) and receptor rnteracting protein kinase 1 (RIP1) leading to the activation of the canonical pathway for NF-κB activation (17~21).
Apoptosis signal-regulating kinase 1 (ASK1) is a MAPK kinase kinase (MAP3K) that is activated in response to cytotoxic stresses, and proinflammatory cytokines such as interlueukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), lipopolysaccharide (LPS), reactive oxygen species (ROS), endoplasmic reticulum (ER) stress and calcium influx (16, 22, 23). In response to these stimuli, ASK1 activates p38 MAPK and JNK, thereby regulating various biological responses which include cell death by apoptosis, inflammation, differentiation, and survival (24).
EBV-positive Burkitt's lymphoma (BL) cells exhibit elevated levels of ROS (25), and the expression of EBV nuclear antigen 2 (EBNA2) or LMP1 alone can induce ROS generation (25). Interestingly, ROS is essential for NF-κB activation, but not for MAPK activation, in EBV-positive BL cells (25). Therefore, in the present study, we investigated the role of ASK1 in EBV LMP1-induced NF-κB activation.
MATERIALS AND METHODS
Cells, plasmids, transfection and reporter gene assays
Maintenance and propagation of HEK293, HeLa, EBV-negative Burkitt's lymphoma BL41, and BL41 cells infected with the B958 strain of EBV (BL41-B958) have been described previously (13, 26). ASK1 knock-out (KO) mouse embryonic fibroblasts (MEFs), pCDNA3-HA-ASK1 (WT), and pCDNA3-HA-ASK1KM (K709M) were kindly provided by Dr. Hidenori Ichijo (The University of Tokyo). pSG5-FLAG-LMP1 was previously described (27). For transient transfection, Effectene transfection reagents were used according to the manufacturer's directions (Qiagen, Valencia, CA, USA). Luciferase assays were performed as previously described (28). Luciferase data shown here represent the mean values of three independent experiments.
Western blot analysis
Cells were collected, fractionated, and transferred to nitrocellulose membranes as described previously (29). Antibodies to HA, ASK1, IRAK1, and TRAF6 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibody to phospho-ASK1 Thr845 was purchased from Cell Signaling Technology (Beverly, MA, USA). Antibody to alpha-tubulin was purchased from Sigma Aldrich (St. Louis, MO, USA). Enhanced chemiluminescence detection reagents (Pierce, Rockford, IL, USA) and secondary peroxidase-labeled anti-mouse or anti-rabbit immunoglobulin G antibody (Amersham Biosciences, Piscataway, NJ, USA) were used according to the manufacturer's directions.
Immunoprecipitation
Ten million cells were lysed in buffer containing 20 mM Tris-HCl [pH 7.5], 100 mM NaCl, 0.5 mM EDTA, 1% NP-40, phosphatase inhibitor cocktail (Calbiochem, La Jolla, CA, USA), and protease inhibitor cocktail (Roche, Indianapolis, IN). Lysates were pre-cleared with protein A/G agarose beads (Santa Cruz Biotechnology) and incubated at 4℃ for overnight with anti-TRAF6 antibody (Santa Cruz Biotechnology). After washing three times with lysis buffer, protein complexes were analyzed by Western blot.
RESULTS
ASK1 is highly activated in EBV-positive BL cells
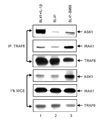
Since EBV-positive BL cell was associated with high levels of ROS (25), the status of ASK1 expression and activation in EBV-positive BL41-B958 cells was determined (Fig. 1). As previously reported (13), the expression of ASK1 was strongly induced in EBV-positive BL41-B958 cells compared to EBV-negative BL41 cells (Fig. 1, compare lane 2 with lane 1). In addition, phosphorylation of threonine 845 in the activation loop of ASK1 was significantly increased in BL41-B958 cells (Fig. 1, compare lane 2 with lane 1). These data indicate that ASK1 is highly expressed and activated in EBV-positive BL41-B958 cells.
The interaction between TRAF6 and IRAK1 or ASK1 is increased in EBV-positive BL cells
Since ROS induced the interaction between TRAF6 and ASK1 (16), we further determined the interaction of TRAF6 to ASK1 in EBV-positive BL cell lines. The lysates of BL41 cells, BL41 cells treated with IL-1β, or BL41-B958 cells were immunoprecipitated with anti-TRAF6 antibody, and ASK1 binding was assessed by Western blot (Fig. 2). In BL41 cells treated with IL-1β or BL41-B958 cells, the interaction between TRAF6 and ASK1 was significantly increased (Fig. 2, compare lanes 1 and 3 with lane 2). In addition, TRAF6 interaction with IRAK1 was also higher in BL41 cells treated with IL-1β or BL41-B958 cells (Fig. 2, compare lanes 1 and 3 with lane 2). Thus, the interaction between TRAF6 and ASK1 or IRAK1 is strongly induced in EBV-positive BL41-B958 cells compared to EBV-negative BL41 cells.
ASK1 is activated by EBV LMP1
Since LMP1 induces ROS (25), we further investigated the role of LMP1 in ASK1 activation (Fig. 3). HeLa cells were transfected with either pSG5 or pSG5-FLAG-LMP1 plus pcDNA3-HA-ASK1, and cell lysates were subjected to Western blot analysis with anti-phospho-ASK1 Thr845 antibody, anti-HA antibody, or anti-LMP1 antibody at 24 h after transfection. Interestingly, ASK1 phosphorylation at threonine 845 was strongly induced in cells transfected with pSG5-FLAG-LMP1, but not in cells transfected with pSG5 (Fig. 3, compare lane 2 with lane 1). These data indicate that LMP1 activates ASK1 in HeLa cells.
ASK1 is required for LMP1-induced NF-κB activation
ROS inhibition significantly reduces NF-κB activation in EBV-positive BL cells (25), and ASK1 has been reported to play an important role in NF-κB activation (14, 15). To assess the role of ASK1 in LMP1-induced NF-κB activation, a dominant negative ASK1 mutant (ASK1KM) was overexpressed in HEK293 cells, and NF-κB activation was determined using an NF-κB-dependent luciferase reporter assay (Fig. 4A). Interestingly, increasing amounts of ASK1KM strongly interfered with LMP1-induced NF-κB activation (Fig. 4A, compare lane 2 with lanes 4 and 6). To further investigate the role of ASK1 in LMP1-induced NF-κB activation, ASK1 wild-type (ASK1+/+) or knock-out (ASK1-/-) MEFs were transfected with either pSG5 or pSG5-FLAG-LMP1, and NF-κB activation was determined using an NF-κB-dependent luciferase reporter assay (Fig. 4B). In ASK1 knock-out MEFs, LMP1-induced NF-κB activation was reduced by 75% compared to ASK1 wild-type MEFs (Fig. 4B, compare lane 4 with lane 2). These data indicate that ASK1 plays a significant role in LMP1-induced NF-κB activation.
DISCUSSION
LMP1 is essential for EBV-infected B lymphocyte transformation to proliferating LCLs and functionally mimics CD40, a member of the tumor necrosis factor receptor (TNFR) superfamily (1). Since LMP1-induced NF-κB activation is critical for EBV-transformed LCL survival (12, 13), delineation of LMP1-induced NF-κB activation pathway may provide a potential drug target in EBV-associated cancers.
Recently, it was reported that the levels of ROS are significantly elevated in EBV-positive BL cells, and ROS is not essential for MAPK activation but contributes to NF-κB activation (25). Since LMP1 alone can induce ROS generation (25), we investigated whether LMP1 activates ASK1 and whether ASK1 is critical for LMP1-induced NF-κB activation.
The data presented here indicate that ASK1 is highly expressed and activated in EBV-positive BL cells, and LMP1 alone can activate ASK1 in HeLa cells. ROS activates TRAF6-ASK1-p38 MAPK pathway that is essential for Toll-like receptor 4 (TLR4) mediated innate immune responses (16). Similar to the TLR4 signaling pathway, TRAF6 is strongly associated with ASK1 or IRAK1 in EBV-positive BL cells. Although there were several reports indicating that ASK1 may be important for NF-κB activation (14, 15), ASK1 has been thought to be essential for p38 MAPK and JNK, but not for NF-κB activation. However, our experiments using both dominant negative ASK1 mutants and ASK1 knock-out MEFs provide strong evidence that ASK1 is required for LMP1-induced NF-κB activation.
Taken together, the results of this study demonstrate that ASK1 is activated by LMP1 and is critical for LMP1-induced NF-κB activation. ASK1 may also play an important role in LMP1-induced p38 MAPK or JNK activation. How LMP1 activates ASK1 and whether ROS or ASK1 is essential for LCL survival are unclear and are the subjects of future studies.



 XML Download
XML Download