

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Foot and mouth disease (FMD) is a devastating disease in cattle, swine, sheep, goats, and many wild cloven-hoofed animal species. The causative agent of the disease belongs to the Aphthoviruses genus of the Picornaviridae family and has a positive sense, single-stranded RNA genome. This disease is characterized by temporary and debilitating oral and pedal vesicles (1).
Despite the success of the inactivated vaccine development, a significant portion of FMD research has been focused on the development of novel vaccines which could be manufactured without the concern of virus mutation or the need for high-containment facilities (2). Disease-free countries hesitate to use vaccination to eliminate FMD because the current vaccine contains varying degrees of contaminating viral non-structural (NS) proteins that make it difficult to differentiate infected from vaccinated animals. In addition, the current vaccine requires 7~14 days for protection to fully develop. This concern is particularly relevant to the control of FMD since FMDV replicates and spreads so rapidly. Prior to induction of the adaptive immune response, animals are still susceptible to new infection. As a result, more rapidly acting vaccines are needed. A number of different anti-viral strategies such as interferon, small interference RNA (siRNA) or single chain fragment of antibody (ScFv) have been tested (3~5).
Interferon (IFN) α/β are cytokines produced for a powerful defense against virus infection. The virus-infected cells express IFN and rapidly achieve an anti-viral state in vivo (6). Type I IFN inhibits replication of picornavirus, retrovirus, influenza virus, vesicular stomatitis virus (VSV), herpes simplex virus (HSV), vaccinia virus, adenovirus and reovirus in vitro (7). Interleukin-18 (IL-18) plays an important role in the cell-mediated immune response, in particular through its ability to induce the production of IFN (8).
Recent pharmacokinetic studies indicate that IFN exhibits an extremely short half-life in the blood after parenteral administration. The poor performance of IFNs may have been caused by an insufficient delivery or lack of sustained delivery of the protein to the active site (9). Recently, delivery systems using adenovirus have been recommended as an effective therapeutic strategy to deliver genes and to express foreign protein in mammals (10~12). The use of adenovirus can induce mucosal immunity and stimulate an immune response in the respiratory and digestive systems (13, 14). Furthermore, the advantages of the adenovirus expression system are its ability to infect various cell types and to deliver foreign genes for expression in host animals like cattle and pigs (15). Adenovirus containing porcine IFNα (Ad-PIFNα) has been reported to be effective against FMDV infection (16). It was well known that porcine IFNα had better anti-FMDV effect than IFNβ (4). Furthermore, bovine IFNα also had anti-FMDV effect (17).
In this study, to induce an antiviral state in vivo, we used adenoviruses expressing IFNα or β. Anti-FMDV effects of human IFN has been questionable in porcine or bovine-originated cells and animals. Therefore, we investigated the potential anti-FMDV effect of human IFN as the first line of defense in the animals. In addition, an adenovirus expressing porcine IFNβ and IL-18 was tested to cover the host-specific response for pig with use of human interferon. IL-18, a known inducer of IFNγ, was used for an additional effect by sustaining the delayed anti-viral state. The fused form for porcine cytokines would be effective as a defense strategy for one shot vaccine carrying function for double effects. Cattle and pig cells, in vitro, and sucking mice were infected with the recombinant adenoviruses for the measurement of their antiviral activity and effect on FMDV replication.
MATERIALS AND METHODS
Virus and cells
Foot and mouth disease (FMD) viruses were isolated by the Animal, Plant and Fisheries Quarantine and Inspection Agency (QIA) in Korea in 2000 and 2002 (O/SKR/2000 or O/SKR/2002). Other FMDV serotypes, O/UKG, Asia1/ISR and A22/Iraq were used for inhibition test of FMDV replication. The viruses were tested in a Bio-safety level (BSL) 3 facility. Human embryonic kidney (HEK) 293A cells were purchased from Q-BIO gene (Carlsbad, CA, USA). For inhibition test in various cell types, primary bovine thyroid and pig-originated IBRS-2 cells were used.
Manipulation of anti-viral genes
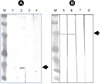
The human IFNalpha (HIFNα) gene was obtained from Cytokine Bank (Chonbuk National University, Korea). The forward (F) and reverse (R) primers for cloning of human IFNα were 5'-GCTCCGGATCCGCTAGCATGTGTGATCTCCCTGAGACCC-3' and 5'-CGTCCGGATCCTTATTCCTTCCTCCTTAATCTTTCTTGC-3', respectively. A BamH I restriction endonuclease site was added to 5' end of the primers for cloning into the expression vector. To obtain porcine IFNβ and IL-18 genes, porcine blood was stimulated with lipopolysaccharide (Sigma-Aldrich, St. Louis, MO, USA). Porcine IFNβ and IL-18 mRNA were extracted and amplified by RT-PCR with IFNβ (IFNβF: 5'-GCTCCGGATCCATGAGCTATGATGTGCTTCGAT-3', IFNβR: 5'-CGTCCGGTACCGTTCCGGAGGTAATCTGTAAGTCTGTT-3') and IL-18 (IL-18F: 5'-CGTCCGGATCCATGTACTTTGGCAAGCTTGAACCTAAAC-3', IL-18R: 5'-GCTCCGGATCCGTTCTTGTTTTGAACAGTGAACATTA-3') specific primers designed in reference to Genbank accession M86762 for porcine IFNβ and AY-450287 for porcine IL-18. PCR was applied with Taq PCR Master Mix (QIAGEN, Hilden, Germany) and the PCR products were cloned to pGEM-T easy vector (Promega, Madison, WI, USA). After insertion of IL-18 gene using Kpn I-linker sequence to the IFNβ-cloned vector for fused gene, the PCR with the primers IFNβF and IL-18R for the fused form was done using a general protocol with an annealing temperature of 56℃. The amplified PCR products of HIFNα and PIFNβ-IL18 were purified by Gene clean III kit (Q-BIO gene, CA, USA). pQBI-AdBM5-GFP (Q-BIO gene) was used as an adenovirus transfer vector by ligation with BamH I-treated PCR products into Bgl II site of the vector (Fig. 1). The HIFNα or PIFNβ-IL18 genes were inserted into cloning site under the control of major late promoter (MLP) of the transfer vector carrying green fluorescent protein (GFP) gene under the control of CMV promoter (pHIFNα, pPIFNβ-IL18). Recombinant adenoviruses were produced in E1 protein expressing HEK 293A cells by Adeno-Quest System (Q-BIO gene). Using fluorescence microscopy, protein expression was examined by fluorescence light emitted from HEK 293A cells.
ELISA to determine an Anti-FMDV activity by antiviral gene from plasmid
Serial diluted DNAs of pHIFNα and pPIFNβ-IL18 were transfected using lipofectamine and plus reagent (Invitrogen, NY, USA) in IBRS-2 cells. At 24 h after transfection, cells were infected with FMDV (100 TCID50/0.1 ml) for challenge. Cytopathic effect (CPE) was examined for 4 days, and then the supernatant from infected cells was harvested and assayed for FMDV antigen ELISA. Anti-FMDV monoclonal antibody was coated on the 96 well ELISA plate for 1 h with coating buffer. After the incubation, the plate was washed with PBS-T (PBS with 0.05% tween 20) three times, and blocked with PBS-T containing 5% skim milk for 1 h. The infected cell supernatant was added to the plate and the plate was placed at 37℃ for 1 h. After the incubation, anti-FMDV MAb HRP conjugate (3H4) was added and the reaction was incubated for another hour. A 3,3_,5,5_-tetramethylbenzidine substrate (TMB, Moss Inc., CA, USA) for colorization was incubated for 15 minute, and optical density was examined within 15 min at 450 nm using ELISA reader (Tecan, Männedorf, Switzerland).
Construction of antiviral protein expressing recombinant adenoviruses
Five micrograms of adenoviral DNA (Q-BIO gene, CA, USA) and EcoR I-linearized pHIFNα and pPIFNβ-IL18 DNA were co-transfected by co-precipitation using calcium phosphate in HEK 293A cells. After 24 h, co-transfected cells were transferred to 6 well plates and overlaid with 2 ml of 1.25% SeaPlaque® agarose (Cambrex Bio Science, NJ, USA) containing 2% FBS in DMEM. Within 2 weeks after co-transfection, the recombinant virus was collected from virus plaque and selected by GFP expression. The viruses were re-infected in HEK 293A cells in 175 cm2 culture dishes. When the cells exhibited CPE of 90%, the cells were freeze-thawed three times, and the stock virus was harvested. The virus was amplified upto the 5th passage to obtain a high viral titer.
SDS-PAGE and Western blot analysis
For protein analysis, recombinant virus infected HEK 293A cells were prepared in lysis buffer and then disrupted by sonication. The clarified supernatant after centrifugation at 13,000 rpm for 10 min was used for 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The electrophoresed proteins in a 12% SDS-PAGE gel were transferred to nitrocellulose membrane. Sheep anti-human IFNα (Biosource, NY, USA) or sheep anti-porcine IFNβ (Biosource) was used as a primary antibody for Western blotting. A HRP-rabbit anti-sheep IgG (H + L) (Zymed, NY, USA) was used for secondary detection with a DAB Substrate kit (Pierce, IL, USA).
Anti-FMDV effect in the porcine or bovine cell infected with recombinant adenovirus
Porcine originated IBRS-2 or bovine originated bovine thyroid cells were pre-cultured for 24 h in 96 well plates.
Recombinant adenoviruses, Ad-HIFNα and Ad-PIFNβ-IL18, were used to infect the cultured cells by 10-fold dilution. At 24 h after infection, the cells were infected superinfected with FMDV (100 TCID50/0.1 ml) for challenge. CPE was monitored for 7 days, after that period the supernatant from infected cells was assayed using a FMDV antigen ELISA for assessing for antiviral effect. FMDV antigen ELISA was determined from the supernatant for FMDV growth at 72 h after FMDV infection. Cells were infected with Ad-HIFNα or Ad-PIFNβ-IL18 (104.0 TCID50/0.1 ml) and FMDV serotypes (102.0 TCID50/0.1 ml) of O/UKG, Asia1/ISR and A22/Iraq were used to challenge cells at 24 h after infection with recombinant adenoviruses. The control group was inoculated with adenovirus expressing lamin-shRNA (Ad-control) (Invitrogen, NY, USA).
Challenge test against FMDV in suckling mice
All experimental animals were cared for according to the animal management guideline of QIA in Korea. Ten of 7 day-old suckling ICR mice per group were injected with Ad-HIFNα (106.5 TCID50/0.1 ml) and Ad-PIFNβ-IL18 (107.5 TCID50/0.1 ml). After 24 h, mice were injected with FMDV (10 to 125 LD50/0.1 ml) for challenge and their survival rate was observed for following 10 days post-challenge.
RESULTS
Effect of IFNα and IFNβ on FMDV replication
IBRS-2 cells were transfected with plasmids containing the antiviral genes HIFNα and PIFNβ-IL18 at various concentrations for comparison of antiviral effects. After 24 h, the cells were infected by FMDV (100 TCID50/0.1 ml). After 48 h, the supernatant was harvested for FMDV antigen quantitation. The plasmid containing HIFNα gene (pHIFNα) showed a better anti-FMDV effect than pPIFNβ-PIL18 (Fig. 2). The lower concentrations of pPIFNβ-IL18 had very little effect on FMDV replication, but a high concentration (1 µg) had a measurable antiviral effect. The plasmid mixture of pHIFNα and pPIFNβ-IL18 showed antiviral activity at the same level as pHIFNα. Both plasmids at a concentration of 1 or 2 µg showed strong anti-FMDV activities, but the pPIFNβ-IL18 at concentration of 3.9 ng to 0.5 µg could not inhibit FMDV replication (Fig. 2).
Construction of recombinant adenoviruses expressing IFNs and IL-18
The molecular weight of proteins expressed in HEK 293A cells was 25 kDa for Ad-HIFNα, and 53 kDa for Ad-PIFNβ-IL18, as determined by Western blot assay (Fig. 3).
To identify inhibition of FMDV replication after recombinant adenovirus infection, FMDV antigen ELISA was performed using the supernatant collected 48 h after FMDV challenge following the adenovirus infection for 24 h in bovine thyroid or IBRS-2 cells. Ad-HIFNα showed strong antiviral effect as demonstrated using plasmid transfection (Fig. 4). The contribution of the PIFNβ-IL18 appeared to be negligible.
Anti-FMDV effect in vivo
Suckling mice were infected with either Ad-HIFNα or Ad-PIFNβ-IL18. As shown in Fig. 5, the group with recombinant adenovirus had a longer survival rate compared to the control group. The group of Ad-HIFNα or Ad-PIFNβ-IL18 showed extended survival of 2 days compared with control group (Fig. 5 and Fig 6). Among the groups, the group infected with Ad-HIFNα had the highest survival rate when inoculated repeatedly.
DISCUSSION
The action of IFN is the first line of defense mechanism to protect against viral infection. The activated cell secretes IFN which binds to the cell receptor, and remains in antiviral states at broad regions. Human adenovirus type 5 has been proven as an effective vector for expression of foreign protein and gene delivery to induce humoral or cellular immunity in digestive and respiratory organs in animals (10~15).
The epithelial surface in oropharyngeal region is favored for entry by many kinds of viruses, bacteria and parasites. Immune responses in mucosal region are an important defense mechanism to protect against the first infection of microorganism. The first replication site of FMDV is an epithelial surface in oropharyngeal region, which is unavailable for the conventional inactivated vaccine. This disadvantage could be overcome by introduction of antiviral molecules into the oropharyngeal regions and could provide a safer tool to protect against FMDV infection.
It has been shown that the pigs infected by adenovirus expressing porcine IFNα by intramuscular (IM) route were protected after FMDV challenge by the same route (16).
In this study, human IFNα showed anti-FMDV activity. Human IFN could be used in many different species that are susceptible to FMDV infection, including pig, cattle and other target species. However, previous study strategies have utilized porcine IFN which is limited to pigs (4). For pigs as a target animal, porcine IFNβ/IL-18 (PIFNβ-IL18) was constructed additionally. Anti-FMDV activity in IBRS-2 cell and suckling mice was assayed by expression of HIFNα and PIFNβ-IL18. We proved that the strategy using HIFNα elicited a better antiviral effect independent of recombinant adenoviral titers. pPIFNβ-IL18 showed anti-FMDV effect only at high concentrations in IBRS-2 cells, and the groups inoculated with Ad-PIFNβ-IL18 showed low survival rates in suckling mice. Using an inhibition test of FMDV replication with the various cell types, an anti-FMDV effect in both bovine and porcine-originated cells was shown (Fig. 4). The results might also demonstrate protection in vivo of susceptible animals like cattle or pigs if a similar mechanism is required. Furthermore, we found the possibility for enhancing the survival of FMDV infected mice by repeat injection until a maximum of 4 injections of the recombinant adenovirus. This phenomenon shows the relationship with the number of injections and duration of the gene expression in vivo because of the short half life of IFN.
To overcome the many disadvantages of the conventional inactivated FMD vaccine, we constructed recombinant adenovirus delivering HIFNα which exhibited an anti-FMDV activity. The constructs are expected to protect transmission of FMDV in replication sites as well as early infection in various viral diseases by the same mechanism of enhancing non-specific innate immunity. A further experiment will be carried out to establish if human IFN works in target animals such as cattle and pigs.




 XML Download
XML Download