

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Bacillus anthracis is a gram-positive, aerobic, spore forming, and rod-shaped bacterium. Anthrax spores are introduced into macrophage by phagocytosis and multiply after germination. The anthrax spores infected in macrophage produce lethal toxin eventually caused cell death. In this study, we analyzed apoptosis and cytokine TNF-α and IL-12 secretion after the infection of spores of B. anthracis Sterne in the murine macrophage RAW264.7 cells and in the primary human macrophages. In murine macrophage RAW264.7 cells infected by spore of B. anthracis Sterne, the cells were markedly changed in secretion of TNF-α (482∼6,213 pg/ml) by lethal toxin, and induced apoptosis. In case of RAW264.7 cells infected by formalin-inactivated spores of B. anthracis, the cells were not able to produce lethal toxin, which released lower level concentration of TNF-α (7.7∼97.2 pg/ml), and rarely induced apoptosis. When primary human macrophage cells infected with spores of B. anthracis Sterne, they secreted TNF-α (5∼16 pg/ml), and induced apoptosis about 1% of total cells. We presented that inducing apoptosis by spores of B. anthracis Sterne capable of expressing lethal toxin is related with the secretion of TNF-α in murine macrophage RAW264.7 cells. These studies revealed that human and murine macrophages has affected differently by anthrax lethal toxin produced by spores of B. anthracis Sterne.
Go to : 
References
1). Agrawal A, Lingappa J, Leppla SH, Agrawal S, Jabbar A, Quinn C, Pulendran B. Impairment of dendritic cells and adaptive immunity by anthrax lethal toxin. Nature. 424:329–334. 2003.

2). Beauregard KE, Collier RJ, Swanson JA. Proteolytic activation of receptor-bound anthrax protective antigen on macrophages promotes its internalization. Cell Microbiol. 2:251–258. 2000.
3). Cerrtti D, Hollingsworth LT, Kozolosky CJ, Valentine MB, Shaprio DN, Morris SW, Nelson N. Molecular characterization of the gene for human interleukin-1-beta converting enzyme (IL1BC). Genomics. 20:468–473. 1994.
4). Chopra AP, Boone SA, Liang X, Duesbery NA. Anthrax lethal factor proteolysis and inactivation of MAPK kinase. J Biol Chem. 278:9402–9406. 2003.
5). Cordoba-Rodriguez R, Fang H, Lankford CS, Frucht DM. Anthrax lethal toxin rapidly activates caspase-1/ICE and induces extracellular release of interleukin (IL)-1beta and IL-18. J Biol Chem. 279:20563–20566. 2004.
6). Dragon DC, Rennie RP. Evaluation of spore extraction and purification methods for selective recovery of viable Bacillus anthracis spores. Lett Appl Microbiol. 33:100–105. 2001.
7). Erwin JL, DaSilva LM, Bavari S, Little SF, Friedlander AM, Chanh TC. Macrophage-derived cell lines do not express proinflammatory cytokines after exposure to Bacillus anthracis lethal toxin. Infect Immun. 69:1175–1177. 2001.
8). Fouet A, Mesnage S. Bacillus anthracis cell envelope components. Curr Top Microbiol Immunol. 271:87–113. 2002.
9). Fritz DL, Jaax NK, Lawrence WB, Davis KJ, Pitt MLM, Ezzell J, Friedlander A. Pathology of experimental inhalation anthrax in the rhesus monkey. Lab Investig. 73:691–702. 1995.
10). Guidi-Rontani C, Mock M. Macrophage interactions. Curr Top Microbiol Immunol. 271:115–141. 2002.
11). Hanna PC, Acosta D, Collier RJ. On the role of macrophages in anthrax. Proc Natl Acad Sci USA. 90:10198–10200. 1993.
12). Jung KH, Seo GM, Kim SJ, Kim JC, Oh SM, Oh KG, Chai YG. Proteome profiling of murine macrophages treated with the anthrax lethal toxin. Kor J Microbiol. 41:262–168. 2005.
13). Kim SJ, Cho KS, Choi YK, Chai YG. Production of Bacillus anthracis protective antigen by improvement of culture condition and purification methods. Kor J Microbiol. 37:1–27. 2001.
14). Klein F, Walker JS, Fitzpatrick DF, Lincoln RE, Mahlandt BG, Jones WIJ, Dobbs JP, Hendrix KJ. Pathophysiology of anthrax. J Infect Dis. 116:123–138. 1966.
15). Krantz BA, Melnyk RA, Zhang S, Juris SJ, Lacy DB, Wu Z, Finkelstein A, Collier RJ. A phenylalanine clamp catalyzes protein translocation through the anthrax toxin pore. Science. 309:777–81. 2005.
16). Klimpel KR, Arora N, Leppla SH. Anthrax toxin lethal factor contains a zinc metalloprotease consensus sequence which is required for lethal toxin activity. Mol Microbiol. 13:1093–100. 1994.
17). Lacy DB, Collier RJ. Structure and function of anthrax toxin. Curr Top Microbiol Immunol. 271:61–85. 2002.
18). Ma X. TNF-αlpha and IL-12: a balancing act in macrophage functioning. Microbes Infect. 3:121–129. 2001.
19). McHugh SL, Yamamoto Y, Klein TW, Friedman H. Murine macrophages differentially produce proinflammatory cytokines after infection with virulent vs. avirulent Legionella pneuphila. J Exp Med. 190:953–962. 2000.
20). Milne JC, Furlong D, Hanna PC, Collier RJ. Anthrax protective antigen forms oligomers during intoxication of mammalian cells. J Biol Chem. 269:20607–20612. 1994.
21). Moayeri B, Haines D, Young HA, Leppla SH. Bacillus anthracis lethal toxin induces TNF-α-independent hypoxia-mediated toxicity in mice. J Clin Invest. 112:670–682. 2003.
22). Mock M, Fouet A. Anthrax. Annu Rev Microbiol. 55:647–671. 2001.
23). Pickering AK, Merkel TJ. Macrophages release tumor necrosis factor alpha and interleukin-12 in response to intracellular Bacillus anthracis spores. Infect Immun. 72:3069–3072. 2004.
24). Pickering AK, Osorio M, Lee GM, Grippe VK, Bray M, Merkel TJ. Cytokine response to infection with Bacillus anthracis spores. Infect Immun. 72:6382–6389. 2004.
25). Popov SG, Villasmil R, Bernardi J, Grene E, Cardwell J, Popova T, Wu A, Alibek D, Bailey C, Alibek K. Effect of Bacillus anthracis lethal toxin on human peripheral blood mononuclear cells. FEBS Lett. 527:211–215. 2002.
26). Popov SG, Popova TG, Grene E, Klotz F, Cardwell J, Bradburne C, Jama Y, Maland M, Wells J, Nalca A, Voss T, Bailey C, Alibek K. Systemic cytokine response in murine anthrax. Cell Microbiol. 6:225–233. 2004.
27). Seo GM, Kim SJ, Chai YG. Rapid profiling of the infection of Bacillus anthracis on human macrophages using SELDI-TOF mas spectroscopy. Biochem Biophys Res Commun. 325:1236–1239. 2004.
28). Smith H. Discovery of the anthrax toxin: the beginning of in vivo studies on pathogenic bacteria. Trends Microbiol. 8:199–200. 2000.
29). Smith H, Stoner HB. Anthrax toxic complex. Fed Proc. 26:1554–1557. 1967.
30). Tonello F, Ascenzi P, Montecucco C. The metalloproteolytic activity of the anthrax lethal factor is substrate-inhibited. J Biol Chem. 274:40075–40078. 2003.
31). Tracey KJ, Fong Y, Hesse DG, Manogue KR, Lee AT, Kuo GC, Lowry SF, Cerami A. Anti-cachectin/TNF monoclonal antibodies prevent septic shock during lethal bacteraemia. Nature. 330:662–664. 1987.
32). Vitale G, Bernardi L, Napolitani G, Mock M, Montecucco C. Susceptibility of mitogen-activated protein kinase kinase family members to proteolysis by anthrax lethal factor. Biochem J. 352:739–745. 2000.
33). Wallach D. Cell death induction by TNF: a matter of self control. Trends Biochem Sci. 22:107–109. 1997.
34). Wewers MD. IL-1 beta: an endosomal exit. Proc Natl Acad Sci USA. 101:10241–10242. 2004.
35). Zaucha GM, Pitt LM, Estep J, Ivins BE, Friedlander A. The pathology of experimental anthrax in rabbits exposed by inhalation and subcutaneous inoculation. Arch Pathol Lab Med. 122:982–992. 1998.
Go to :
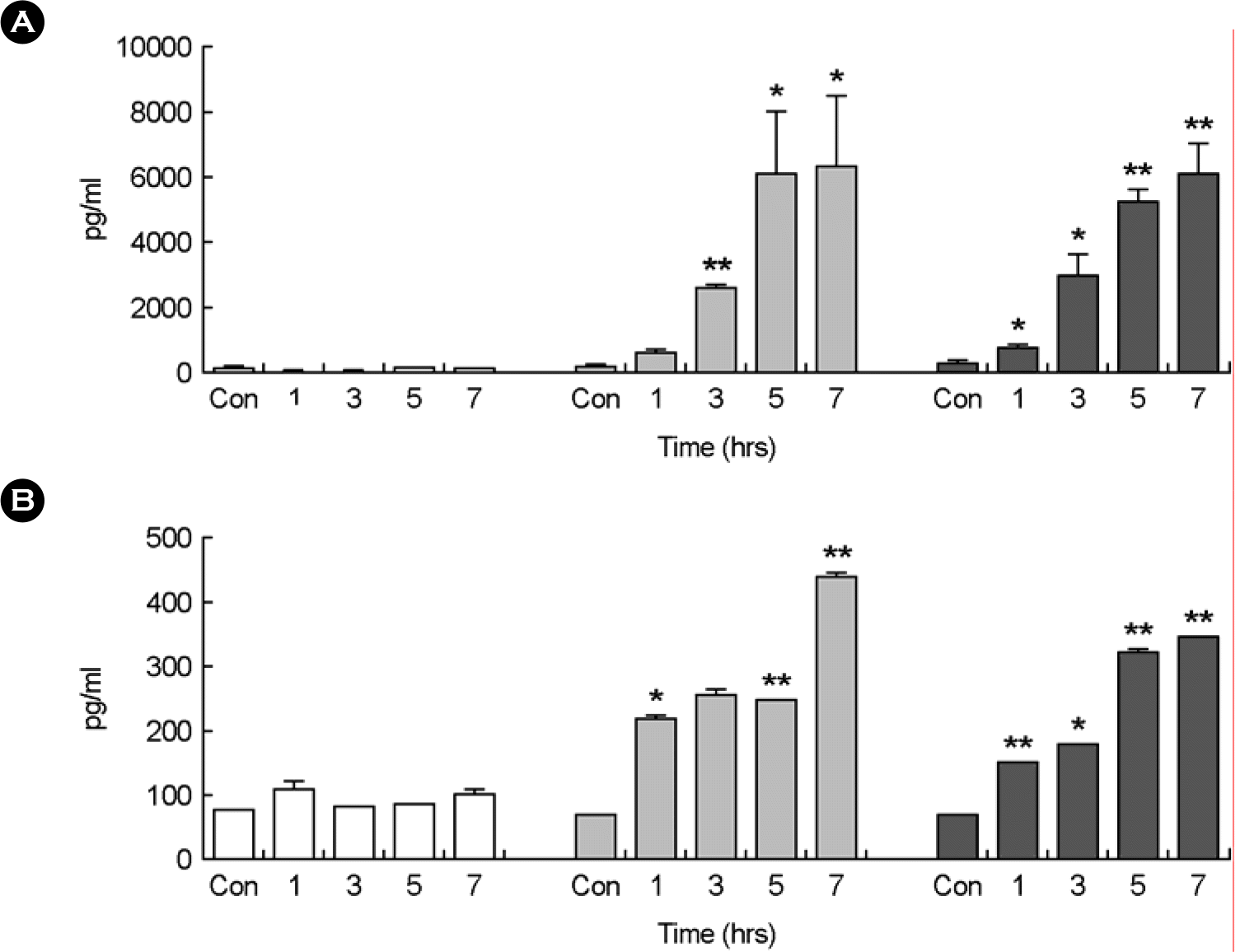 | Figure 1.Cytokine analysis in murine macrophage RAW264.7 cells infected with spores of B. anthracis Sterne at 1, 3, 5, and 7 hours using ELISA method. Control was murine macrophage RAW264.7 cells treated with PBS buffer. The concentration of 2 cytokines in the supernatants collected from murine macrophage RAW264.7 cells infected with formalin-inactivated spores (white bars), spores (gray bars), and vegetative cells (black bars) of B. anthracis Sterne at the indicated time points are shown for TNF-α (A), and IL-12 (B). Three experiments were performed, and the results of a representative experiment are shown. Each value reported is the average of three samples. Error bars represent 1 standard deviation. Statistical significance was determined by Student's t test analysis. In all cases, means were compared to control group only treated with PBS buffer. ∗p<0.05, ∗∗p<0.01 |
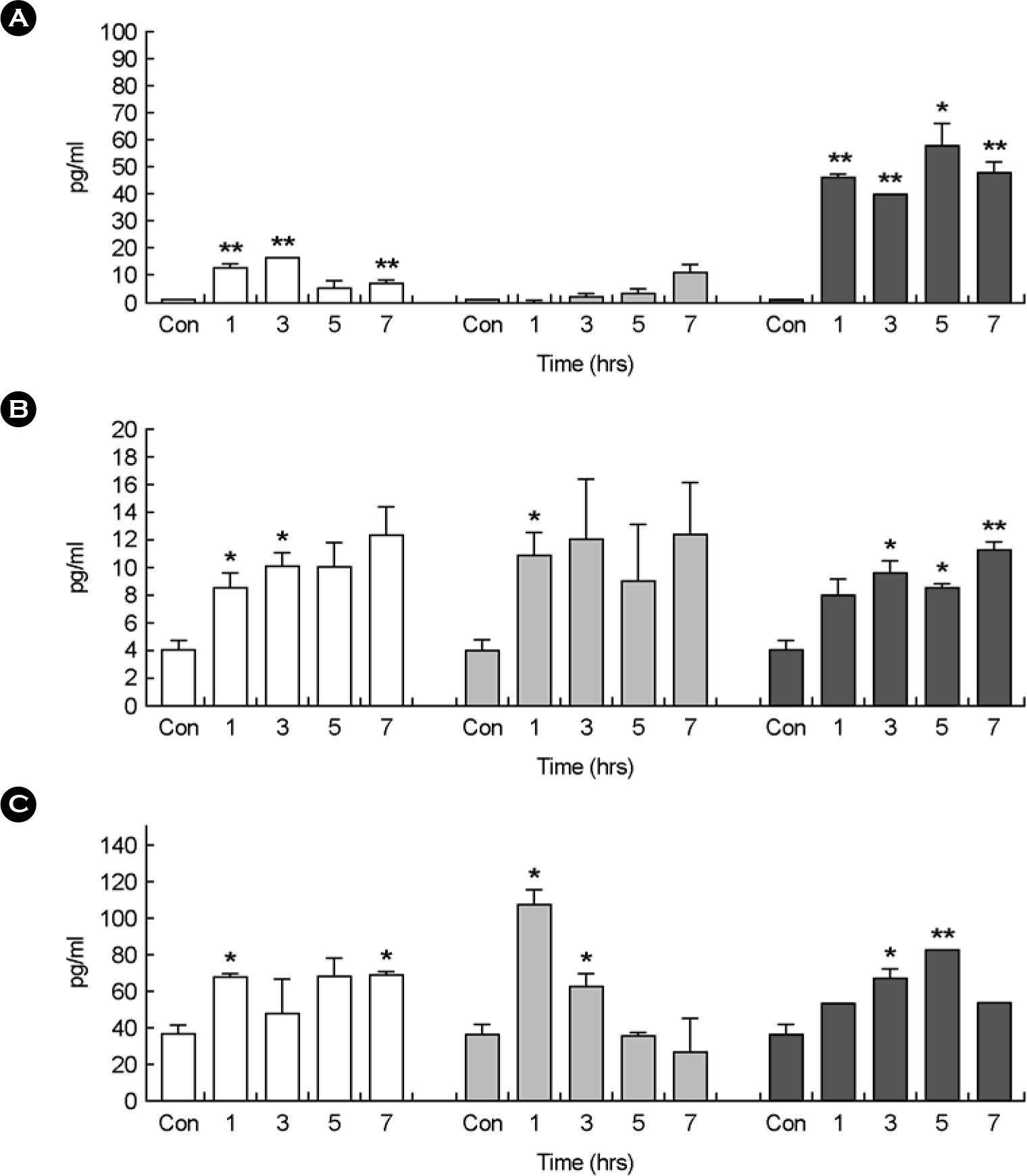 | Figure 2.Cytokine analysis in primary human macrophage infected with spores of B. anthracis Sterne at 1, 3, 5, and 7 hours using ELISA method. Control was murine macrophage RAW264.7 cells treated with PBS buffer. The concentration of 3 cytokines in the supernatants collected from primary human macrophage infected with formalin-inactivated spores (white bars), spores (gray bars), and vegetative cells (black bars) of B. anthracis Sterne at the indicated time points are shown for TNF-β (A), IL-1β (B), and IL-12 (C). Three experiments were performed, and the results of a representative experiment are shown. Each value reported is the average of three samples. Error bars represent 1 standard deviation. Statistical significance was determined by Student's t test analysis. In all cases, means were compared to control group only treated with PBS buffer. ∗p<0.05, ∗∗p<0.01 |
 | Figure 3.Apoptosis analysis in murine macrophage RAW264.7 cells infected with spores of B. anthracis Sterne at 1, 3, 5, and 7 hours by flow cytometry with annexin-PI double-labeled cells. The annexin-positive population corresponding to apoptotic cells, PI-positive population corresponds to necrotic cells, and the double-positive position corresponds to necrotic and/or late apoptotic cells. Murine macrophage RAW264.7 cells were treated with PBS buffer (A), formalin-inactivated spores (B), spores (C), and vegetative cells (D) of B. anthracis Sterne. |
 | Figure 4.Apoptosis analysis in primary human macrophages infected with spores of B. anthracis Sterne at 1, 3, 5, 7 hours by flow cytometry with annexin-PI double-labeled cells. The annexin-positive population corresponding to apoptotic cells, PI-positive population corresponds to necrotic cells, and the double-positive position corresponds to necrotic and/or late apoptotic cells. Primary human macrophages were treated with PBS buffer (A), formalin-inactivated spores (B), spores (C), and vegetative cells (D) of B. anthracis Sterne. |
Table 1.
Cytokine expression by anthrax lethal toxin on different cells of immune system
| Cell Lines | Toxin | Time (hr) | Cytokines | References |
|---|---|---|---|---|
| RAW264.7 | LeTx∗ | 2∼4 | TNF-α ↑ | 11 |
| RAW264.7 | LeTx∗ | 24 | IL-1β, IL-18 ↑ | 5 |
| J774A.1 | LeTx∗ | 2 | TNF-α ↓ | 7 |
| J774A.1 | spore | 5∼7.5 | TNF-α, IL-12 ↑ | 23 |
| Human macrophage | LeTx∗ | 48 | TNF-α ↓, IL-1β, IL-6 ↑ | 25 |
| Mouse macrophage | spore | 5∼7.5 | TNF-α, IL-6 ↑ | 24 |
Table 2.
The amounts of TNF-α or IL-12 secreted from murine macrophage RAW264.7 cells were infected with spores of B. anthracis Sterne
| TNF-α (pg/ml) | IL-12 (pg/ml) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 hr | 3 hr | 5 hr | 7 hr | 1 hr | 3 hr | 5 hr | 7 hr | |
| Inactivated spores | ND∗ | ND∗ | ND∗ | ND∗ | 215 | 254 | 210 | 234 |
| Live spores | 482 | 2,433 | 6,022 | 6,213 | 110 | 246 | 210 | 234 |
| Vegetative cells | 692 | 2,844 | 5,128 | 6,045 | 151 | 175 | 318 | 340 |
 XML Download
XML Download