

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Malignant melanoma is the most aggressive skin cancer due to its capacity to rapidly metastasize and strong resistance to anti-cancer therapy, such as chemotherapy and radiotherapy (1). In spite of extensively researched melanoma physiology, the associated rate of mortality is high and critical therapeutic approaches are still not sufficient. Several reports showed that treatment with antioxidants is more beneficial in preventing the rapid growth and metastasis of melanoma cells (2,3). In particular, the combination of anti-cancer drug and anti-oxidant is more effective than single treatment because the combination treatment leads to overall low toxicity via synergistic effect (4).
Selenium is an essential trace element involved in cellular homeostasis and protection against cellular stress such as ROS (reactive oxygen species) and DNA damage (5). Patrick et al., report that selenium has a function as an anti-cancer agent (6). Several reports demonstrate that sodium selenite, a source of selenium in many food supplements, can strongly induce cell death in various cancer cells (7-9). Additionally, recent studies reveal that sodium selenite display effective anti-cancer activity against melanoma cancer cells. Despite much evidence about the anti-cancer and anti-oxidant effects of selenium, the exact mechanism whereby this chemical induces cell death in various cancer cells is not yet understand.
Recent studies revealed that ROS play a pivotal role in selenium-mediated cancer cell death (10-12). The ROS level increase cell death following treatment with selenium by inducing DNA breakage and cell cycle suppression. It is well known that the CDK (cyclin-dependent kinase) has a critical role in ROS related cell cycle regulation. Some studies report that activation of p53 is induced by CDK2 gene expression upregulated by treatment with selenium, which then induces cell death in mouse mammalian epithelial carcinoma cell line. Moreover, activation of p53 and p38 pathway has been shown to induce apoptosis in human cervical cancer following treatment with selenium (13-16). Although the effects of selenium on the growth and metastasis of some cancer cells are well known, the anti-cancer effects of selenium on cancer cell growth and cell migration in melanoma are not completely understood. In this study, we evaluate the anti-cancer effects of selenium on a murine melanoma cell line and the anti-metastatic ability of selenium on an experimental metastasis model using B16 mouse melanoma cells. These findings provide useful information on the therapeutic tool of antioxidant treatment metastatic melanoma cancer.
MATERIALS AND METHODS
Cell culture and reagents
B16F10, murine melanoma cell line, was maintained in RPMI 1640 supplemented with 2 mM L-glutamine, 100 units/ml penicillin, 100 ug/ml streptomycin and 10% heat-inactivated fetal bovine serum. The cells were cultured at 37℃ in a humidified atmosphere that contained 5% CO2. These cells were used for experiments while they were in the log phase of growth. Sodium selenite (selenium) was purchased from Sigma (St. Louis, MO, USA).
Assay for viable cells
The inhibition of cell growth was determined by 3 (4, 5-dimethyl thiazolyl-2) 2, 5 diphenyl tetrazolium bromide (MTT; Sigma, St. Louis, MO, USA) assay. B16F10 mouse melanoma cells were seeded on 96 well plates (1×103 cells/well) and treated with 0, 1, 2, 10, 25, 50, or 100 uM selenium for 72 hrs before adding 20 ul MTT solution (0.5 mg/ml). The MTT absorbance was measured at 570 nm using an ELISA reader (Bio-Tek instruments, Inc., Winnoski, VT, USA) 4 h after addition of the dye. Each experiment was performed in triplicate.
Detection of selenium-mediated apoptosis
To investigate the apoptosis-inducing effect of selenium, cells were collected, washed twice with PBS, and then resuspended in 100 ul of Annexin V binding buffer (10 mM HEPES/NaOH pH 7.4, 140 mM NaCl, 2.5 mM CaCl2). After 2 ul of FITC-conjugated Annexin V (BD Pharmingen, San Diego, CA, USA) and 2 ul of propidium iodide (BD Pharmingen) were added, cells were incubated in the a dark room at room temperature for 15 minutes with gentle shaking. Finally, 400 ul of Annexin V binding buffer was added to each tube and cells were analyzed using a FACSCalibur (BD Pharmingen).
Cell cycle analysis
Cell cycle analysis was performed using propidium iodide (PI)-staining method. Cells were harvested after selenite treatment, washed twice in PBS (2% FBS), and fixed with 70% cold aqueous ethanol at 4℃ overnight. The cells were resuspended in a PI stain (10 ug/ml RNase A and 10 ug/ml PI in PBS) at room temperature for 30 minutes, and DNA content was determined using a FACSCalibur (BD Pharmingen).
Reverse-transcription polymerase chain reaction
To assess the effect of selenite on cell cycle regulatory gene expression, the cells were harvested after selenite treatment and washed twice in cold PBS. RNA was extracted using an RNeasy mini kit (Quiagen, Valencia, CA) according to the manufacturer's instructions, and RT premix (Bioneer, Daejeon, South Korea) was used to transcribe cDNA. RT-PCR analysis of specific gene transcript was carried out using the following primers for p53 (upstream primer; 5'-ATG ACT GCC ATG GAG GAG T, downstream primer; 5'-CTC GGG TGG CTC ATA AGG TA, 664-bp product), p21Waf1 (upstream primer; 5'-CTT TGA CTT CGT CAC GGA GAC, downstream primer; 5'-AGG CAG CGT GAA GGT GTT TGG GG, 480-bp product), CDK2 (upstream primer; 5'-CAC AGC CGT GGA TAT CTG GAG, downstream primer; 5'-TTG CGA TAA CAG GCT CCG TC, 253-bp product), CDK4 (upstream primer; 5'-ACG CCT GTG GTG GTT ACG CT, downstream primer; 5'-CCA TCT CTG GCA CCA CTG AC, 280-bp product). β-actin (upstream primer; 5'-AAG AGC TAT GAG CTG CCT GA, downstream primer; 5'-CAG GAG GAG CAA TGA TCT TG, 300-bp product) was used as an internal control. PCR (24cycle; 30 seconds at 94℃, 30 seconds at 60℃, 60 seconds at 72℃) was performed using AccuPower PCR premix (Bioneer).
Western blots
Cells (1×107 cells/sample) were scraped into cold PBS, centrifuged and the lysed in RIPA buffer (EIPiS, Daejeon, South Korea) on ice for 30 min. Each cell lysate was separated by 15% SDS-PAGE under reducing conditions and transferred to a nitrocellulose membranes using semidry technique at 80mA for 2 hours. The blots were blocked with 5% skim milk in PBS for 1 hour, and subsequently incubated with primary antibodies (Abs) at a final dilution of 1:1,000 for 2 hour. After three washes in PBST (0.05% tween-20 in PBS), blot were incubated with peroxidase-conjugated secondary antibodies (final dilution, 1:3,000) in dilution buffer (5% skim milk in PBS) for 1 hour and then washed as described above. Detection was performed by chemiluminescence using an ECL-kit (Enhanced Chemi Luminescence; Amersham Life Science, Braunschweig, Germany) and the Multiple Gel-DOC system (Fujifilm, Tokyo, Japan). The following primary antibodies were used: p21 Waf1/Cip1, CDK4, CDK2 and β-actin from Cell Signaling (Beverly, MA, USA).
Animal and experimental metastasis assay
Six- to 8-week-old C57BL/6 mice were obtained from Orient (Seoul, Korea). The institutional review board of animal experiments reviewed all animal studies. Six- to 8-week-old C57BL/6 mice were injected with 0.1 ml of PBS containing 0.1 million B16F10 cells by intra-optic venous route. Mice were randomly assigned to 5 groups (5 mice/group). The mice were treated from day 1 onward with 10 uM selenium through intraperitoneal route. On days 4 and 8 the mice received selenium in 200 µl PBS injected i.p. The mice were killed on day 14, and melanotic colonies on the surface of the lung were counted. The lung sections were fixed, stained with hematotoxylin-eosin (H&E) and was observed under a phase contrast microscope.
RESULTS
Effects of the selenium on B16F10 cell growth and cellular morphology
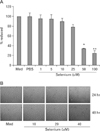
To assess whether selenium directly induces cell death of B16F10 melanoma cells, cells were treated with 0, 1, 2, 10, 25, 50, or 100 uM of selenium. Selenium-induced inhibition of cell growth was determined by MTT assay. Fig. 1A shows that the growth of cells exposed to 50 uM selenium for 72 hour was significantly reduced to 35% of control group. We also found that selenium inhibited the growth of cells in a dose-dependent manner. To confirm our result, we observed the morphological changes of B16F10 cells exposed to 40 uM selenium at 24 hours and 48 hours. As shown in Fig. 1B, treatment with 40 uM selenium significant decreased the cell number of B16F10 melanoma cells in a time-dependent manner.
Effects of the selenium on B16F10 cell death and cell cycle
Next, we measured apoptosis of B16F10 cells by flow-cytometry analysis. Cell cultures were treated under various conditions for 72 hours. Fig. 2A shows that treatment with selenium induces apoptosis in B16F10. The number of apoptotic cells following treatment with 40 uM selenium was significantly higher as compared with the low-dose treated group.
To determine the mechanism of selenium-induced apoptosis, we analyzed the cell cycle of selenium treated B16F10 by PI staining method. Addition of sodium selenium (40 uM) significantly increased the G0/G1 arrest in B16F10 cells compared with the low-dose treated group (Fig. 2B). Taken together, these results indicate that G0/G1 cell cycle arrest is dominantly involved in selenium-induced apoptosis.
Effects of the selenium on expression of cell cycle regulator molecules in B16F10 melanoma cells
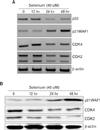
In order to examine the mechanism of selenium-induced G0/G1 arrest in B16F10 cells, we investigated the mRNA and protein expression pattern of various cell cycle regulators such as p53, p21Waf1, CDK2 and CDK4 after treatment with selenium. As shown in Fig. 3A, treatment with selenium significantly decreased expression of p53, CDK2 and CDK4 mRNA in time-dependent manner. However, p21Waf1 mRNA was increased in a time-dependent manner following treatment (Fig. 3A). Furthermore, after exposing cells to 40 uM selenium, the protein level of p21WAF1 significantly increased, while protein levels of CDK2 and CDK4 decreased in time-dependent manners (Fig. 3B). This result implies that selenium-induced cell cycle arrest is regulated by a p53-independent p21Waf1 pathway.
Effects of selenium on tumorigenesis and B16F10 metastasis to mouse lung
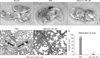
Treatment with selenium regulated p53-independent pathway and cell growth, which are both related to tumor cell growth. To evaluate the effect of selenium on melanoma cell growth, the in vivo growth of melanoma cells in C57BL/6 mouse was determined after the subcutaneous inoculation of B16F10 cells and treatment with selenium. When selenium was administered syngeneic mice, severe regression of tumor growth was not observed (Fig. 4). However, B16F10 melanoma-bearing mice treated with selenium had a significantly reduced tumor burden as measured by counting the number of lung colonies (Fig. 5A). Additionally, we tested for lung metastatic tumor cells to B16F10 bearing mice using histochemical H&E staing. As shown in Fig. 5B and C, treatment with selenium significant decreased the number of lung metastatic spots by B16F10 melanoma cells. These results demonstrate that selenium promotes anti-cancer activity and anti-metastasis activity in melanoma cell bearing mice.
DISCUSSION
One of the most important problems with removing melanoma is its powerful metastatic tumor growth. In the present study, we have evaluated the effect of selenium on controlling B16F10 melanoma proliferation and metastasis. It is well known that anti-oxidant compounds, such as ascorbic acid and selenium, have the ability to inhibit cancer growth and metastasis (2,3). Selenium compound has been specifically reported to significantly reduce cell proliferation and incidence of cancer in vivo by being added to in vitro cell culture or supplemented in animal diets (6). However, the detailed mechanism of its anti-cancer and anti-metastatic effects remains poorly understood.
We evaluated the effects of selenium on the lung metastasis of B16F10 melanoma cells in vivo. Interestingly, our results show that the selenium-treated group had reduced metastasis of B16F10 melanoma cells, although treatment with selenium did not reduce the tumorigenesity of B16F10 melanoma cells (Fig. 4, 5). To identify the affects of selenium on the relationship of tumorigenesity and metastasis in B16F10 melanoma cells, treated cells were analyzed for changes in cell morphology and apoptosis (Fig. 1, 2). Treatment with selenium led to morphologic changes in B16F10 melanoma cells and increase in apoptotic events in a dose-dependent manner. These results suggest that significant anti-metastatic activity of sodium selenite may be due to the apoptotic activity of selenium.
More interestingly, our results revealed that cell apoptosis of B16F10 melanoma cells mediated G0/G1 arrest following treatment with selenium. This is significant because it is well known that regulation of the cancer cell cycle is one of the therapeutic targets for identification of new anti-cancer strategy. Of particular importance are disruptions of cell cycle at the G0/G1 check-point, which leads to dis-regulation of tumor growth (17-21). Based on this evidence, we focused on the mechanism of G0/G1 cell cycle arrest in selenium-induced apoptosis. Many studies have reported that p53 is involved in G0/G1 cell cycle arrest (22-25). However, Deeds et al. recently observed that G0/G1 arrest is induced in a p53-independent manner (26). As shown in Fig. 3, our results support the findings that selenium induces G0/G1 arrest via p53-independent pathway through expression of p21Waf1. Several studies have revealed that p21WAF1 has a critical role in the mechanism of G0/G1 arrest, which involves association of CDK-cyclin complex to inhibit CDK kinase activity (27). We found that treatment of melanoma with sodium selenium induces expression of p21WAF1, which leads to down-regulation of CDK4 and CDK2 expression at the mRNA and protein levels (Fig. 3). Our findings suggest that selenium induces cell apoptosis due to G0/G1 arrest through a p53-independent p21WAF1 pathway.
In conclusion, we suggest that treatment with selenium enhances the ability of anti-metastasis to promote apoptosis of B16F10 melanoma cells by inducing p53-independent G0/G1 cell cycle arrest. Further knowledge regarding the treatment of selenium may provide a usefully concept for therapy strategy to improve malignancy in cancer patients.


 XML Download
XML Download