

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The lymphatic vasculature system is important for at least three functions; first, it controls the absorbance of nutrients including proteins and lipids from blood vasculature into peripheral tissues. Moreover, other small molecules such as cytokines and chemokines of interstitial fluids are transported through the lymphatic vessels. Second, lymphatic vessels are used as a route of immune cell trafficking from peripheral tissues into lymph nodes. Lymphocytes and dendritic cells that have encountered foreign antigens can access the lymphatic capillaries dependent on chemokine gradients (12). Lastly, the lymphatic vasculature system maintains the homeostasis of tissue fluid by regulating lymph transport. The lymph fluid drains into the thoracic lymphatic duct, then gets back into blood circulation (3). This lymphatic transport is tightly regulated by the osmotic gradients and fluid barrier filtration from cytokines, chemokine, and other macromolecules (2). Accordingly, individuals with abnormal lymph circulation or surgery to remove a lymph node can develop lymphedema with impaired immune responses. In this review, we demonstrate the general functions of the lymphatic vasculature system and then discuss studies from the perspective that the lymphatic system is closely linked with the pathophysiology of inflammatory conditions and can control them. Especially, we focus on elucidation of how specialized lymphatic vasculature coordinates multiple physiological roles, how lymphatic vessels regulate immune cell trafficking, and how lymphatic vessels themselves respond to inflammation.
Go to : 
LYMPHATICS IN THE CIRCULATORY SYSTEM
In our body, the vascular circulatory system consists of blood and lymphatic vessels, both of which are lined by endothelial cells. Lymphatic vasculature is composed of a branched network of capillaries and collecting lymphatic vessels. Unlike blood vasculature, lymphatic capillaries are blind-ended and lack pericytes. As an open system, the ubiquitous network of lymphatic capillaries (10 to 80 µm in diameter) and vessels transport interstitial fluid from the arterial ends of blood vessels back into the main circulatory system. Interstitial fluid enters the lymphatic system through single-layered lymph capillaries. The discontinuous basement membrane and weak cell-to-cell attachments make the lymphatic capillaries highly permeable, thereby facilitating uptake of proteins including nutrients, viruses, and other large solutes, which in general have very low diffusivity (4). Unidirectional fluid flow from the capillaries toward the lymph nodes is regulated by lymphatic valves, which inhibit its backflow. The individual capillaries merge to form large lymphatic collecting vessels. The lymphatic collecting vessels are segmented into regions called lymphangions that are interconnected by semi-lunar valves that prevent backflow of the lymph through the lumen of the lymphangions. Synergistic functioning of these two types of valves coupled with rhythmic contraction and relaxation of the smooth muscular layer of the lymphangions produces a motion and forces lymph from one lymphangion to the next. The lymphangions finally merge into afferent lymph vessels, which subsequently drain into the subcapsular sinus of various lymph nodes located throughout the body.
Recent studies suggest that the major source of lymphatic endothelial cells during development in mammals is generated through trans-differentiation from embryonic veins (5). During differentiation into lymphatic endothelial cells (LECs), prospero-related homeodomain transcription factor (Prox1) is essential for lymphaticendothelium- specific programming. Prox1 downregulates blood markers and promotes expression of vascular endothelial growth factor receptor (VEGFR)-3, which is important for LEC survival, migration, and proliferation. LECs then start to bud by following the gradient of a VEGFR-3 ligand, VEGFC (6). As a selective marker for lymph capillary endothelial cells, lymphatic vessel endothelium hyaluronan receptor (LYVE-1) has been utilized commonly to study the lymph vessel network. In addition, to visualize the lymphatic vessels in vivo, Prox1-fluorescently-labeled transgenic animals that expresses GFP or tdTomato driven by the promoter of Prox1, a master regulator in lymphatic development, have been broadly used. Adult lymphatic vessels could be easily detected in peripheral tissue such as the trachea, skin, ear, intestine, and diaphragm (7). Such identification, in recent decades, of several key regulators and markers specific to LECs has accelerated findings that LECs actively interact with other cells.
Go to :
LYMPHATIC VESSELS FROM PERIPHERAL TISSUES TO LYMPH NODES
Together with macrophages and monocytes, dendritic cells (DCs) belong to the mononuclear phagocyte system. Both macrophages and DCs are in close contact with LECs for adaptive immunity and tolerance. Early reports showed that lymph-borne antigens arrive in lymph nodes only a few minutes after subcutaneous injection (8). As a secondary lymphoid organ, a lymph node is the main place to activate an immune response as well as the terminal end of the lymphatic system before the lymph fluid is drained into the circulatory system. Before draining into the vein of the circulatory system, the lymph fluid is screened for foreign antigens in the lymph node through the efferent lymphatic vessels. Strategically positioned along lymphatic vessels of the subcapsular sinus, lymph node-resident macrophages play a key role in filter stations, preventing systemic pathogen dissemination (9). Furthermore, the lymphatic vasculature system supports immune cell migration as well as lymph fluid circulation from a tissue to lymph nodes. Lymphatic capillaries and collecting vessels require differential several steps to move DCs into a lymph node (1). By highly expressing CCL19 and CCL21, lymphatic capillaries attract DCs expressing CC-chemokine receptor 7 (CCR7). In addition to chemokine gradient-dependent migration, it was proposed that their migration is dependent on amoeboid movement rather than integrin-dependent movement in a steady state (210). In a pathological condition, dendritic cells are activated by pathogen-associated molecular patterns (PAMPs) such as Toll-like receptors (TLRs), and their movement depends on integrins such as ICAM-1 and VCAM-1. Once DCs enter the lymphatic capillaries from peripheral tissues, DCs crawl onto LECs toward the lymphatic collecting vessels by taking advantage of binding CCR7 to its ligands CCL19 and CCL21. CCL21 deficiency completely impairs the DC migration, suggesting that intra-lymphatic migration of DCs is dependent on the CCL21 chemotactic gradient (11). Upon arrival into the lymphatic collecting vessels, lymphatic fluid force leads passive transport of DCs into the subcapsular sinus of a lymph node. In lymphatic collecting vessels, the direction and rate of lymphatic flow are regulated by lymphatic valves and lymphatic muscle. Lymphatic muscle surrounding LECs contributes to the contraction of lymphatic collecting vessels by regulating lymph pressure (12). In addition to the lymphatic valve and muscle, DCs surrounding the lymphatic collecting vessels affect the permeability of lymphatic collecting vessels by interacting with LECs. In the absence of CCR7, the interaction between DCs and the endothelial cells of lymphatic collecting vessels is highly reduced because of the loss of CCR7–CCL21 interaction (12). Lymphatic collecting vessels show various phenotypes in CCR7-deficient mice. First, collagen deposition is dramatically increased on the collecting vessel wall, resulting in fibrosis. Second, the permeability of collecting vessels is significantly elevated in CCR7-deficient mice, implying that CCR7+ DCs control lymphatic permeability. Lastly, the removal of a lymph-derived tracer is altered in CCR7-deficient mice, suggesting that lymphatic transport is associated with DCs. In addition to the regulation of lymphatic permeability, lymphatic collecting vessels seem to orchestrate a local immune response by adipose tissue myeloid cells including DCs (13). Injected into skin, fluorescent antigen tracers are delivered to CD11c+ MHCII+ DCs in adipose tissue through the collecting lymphatic vessels, suggesting that these DCs sample lymph-borne antigens during local inflammation and T-cell infiltration (Fig. 1). In addition, CD11b+ Gr1+ myeloid cells suppress the contraction of lymphatic collecting vessels in an inflammatory condition (14). Overall, lymphatic flow in lymphatic collecting vessels is tightly regulated by spatially distinct LECs and by the cross-talk between LECs and tissue-resident DCs. Nonetheless, further research is necessary on the molecular mechanism explaining how peripheral tissue resident DCs regulate LECs.
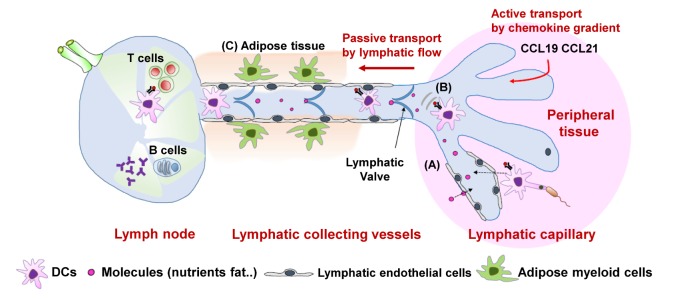 | Figure 1Schematic models of lymphatic flow. (A) Activated DCs can enter the lymphatic capillaries in a CCR7-dependent but integrin-independent manner. Other molecules such as proteins and lipids can be absorbed in lymphatic capillaries. (B) In capillaries, lymphatic endothelial cells drive DCs toward lymphatic collecting vessels by forming chemokine gradients. (C) Once DCs enter the collecting vessels, the interaction of adipose myeloid cells and collecting endothelial cells controls lymphatic flow, which facilitates passive transport of DCs into a lymph node.
|
Go to :
THE ROLE OF LYMPHATICS IN THE REGULATION OF TOLERANCE
As discussed above, the lymphatic vasculature provides routes for DC and lymphocyte migration into and out of a lymph node. LECs control these processes by expression of chemokines and adhesion molecules. LECs also can express immunosuppressive factors such as TGF-β, indoleamine-2,3-dioxygenase, and nitric oxide to maintain peripheral tolerance to self-antigens in lymph (1516) In this regard, LECs participate in the immune response by releasing an appropriate chemokine and cytokines in response to environmental changes in peripheral tissues. Emerging studies demonstrate that lymphatic vessels exhibit tolerogenic potential by regulating T-cell fate and function in immunity. First, it has been shown that LECs can directly dampen DC maturation (17), in turn reducing their ability to activate effector T cells. Second, LECs constitutively express and present a self-antigen on MHC class I molecules to delete autoreactive T cells and thus help maintain peripheral tolerance to these self-antigens (18). LECs can drive deletional tolerance of naïve CD8 T cells without costimulatory molecules necessary to activate effector T cells, and instead express the inhibitory ligand PD-L1 (19). Third, it is now known that LECs can actively scavenge exogenous lymph-borne antigens and process them for cross-presentation on MHC class I molecules (2021). In summary, LECs may potentially help to maintain tolerogenic pressure by presenting both a scavenged and endogenous antigen.
Go to :
THE ROLE OF LYMPHATICS IN INFLAMMATION
The inflammation process is accompanied by several localized or systemic events such as vascular responses, migration and activation of leukocytes, and systemic inflammatory response syndrome. Additionally, lymphangiogenesis frequently occurs during inflammation. To relieve the hostile microenvironment causing inflammation, a harmful antigen and debris should be removed by drainage of excessive interstitial fluid through lymphatic vasculature. Upon initiation of inflammation, the lymphatic system is activated, and both the lymphatic vessels in peripheral tissues and inside a lymph node expand in response to inflammation. Accumulating evidence suggests that inflammation-associated lymphangiogenesis is not an endpoint phenotype of inflammation but rather a type of dynamic and active reaction that can control the inflammatory response or tissue repair (22). Lymphatic vessels newly generated by inflammation play a role in regulation of repair of a damaged tissue (23) and macrophage/DC recruitment, which is critical for pathogen clearance (24). The infiltrating monocytederived macrophages in the lymphatic vessel induce lymphangiogenesis by secreting lymphangiogenic growth factors (VEGF) (25). These ligands (VEGF) secreted from infiltrated macrophages could be major mediators of pathogen clearance and inflammation resolution through profound expansion of lymphatic vessels, enhanced lymph flow, and mobilization of inflammatory cells from the inflamed skin to the draining lymph nodes (26). Such interactions between lymphatic vessels and immune cells contribute to the immune system's response to inflammatory stimuli. Consequently, the inflammationassociated lymphangiogenesis may directly influence the degree of mucosal edema and leukocyte infiltration. Impairment of lymphatic drainage caused by infection, surgery, or trauma results in lymphedema with swelling of limbs or other peripheral mucosal tissues. The associated lymphostasis causes persistence and accumulation of the antigen, foreign material, and immune complexes in the interstitium, which can cause chronic localized inflammation and fibrosis. This chronic inflammation also attenuates lymphatic contraction, hindering lymphatic flow to the lymph node and disrupting the trafficking of lymphocytes to the lymph node. Thus, lymphedema cumulatively results in a region of local immune suppression (27). Accordingly, altered immunity has been reported in lymphedema, highlighting the importance of lymphatic pathways in adaptive immune responses in a variety of pathologies. Overall, the role of lymphatic vessels in the modulation of the immune system in terms of response to inflammation caused by environmental stimulation of peripheral tissues is crucial for homeostasis.
In the detailed molecular mechanism of lymphangiogenesis, pro-lymphangiogenic signaling pathways, among which the VEGF-C and D/VEGFR3 axis is the best characterized, are activated by inflammatory stimuli. Inhibition of the VEGF-C/VEGFR3 axis suppresses lymphangiogenesis and relieves the severity of inflammatory symptoms in rheumatoid arthritis and LPS-induced acute inflammation (26). In addition, VEGF-A either promotes lymphatic vasculature formation via activation of VEGFR2 in repair associated lymphangiogenesis (28). However, T cells negatively regulate lymphatic vessels through T helper 1 (Th1) or Th2 cytokines (anti-lymphangiogenic factors) (29). During the acute stage of inflammation, the pro-lymphangiogenic factors usually dominate, in which case, finally, lymphatic vessels are amplified. As inflammation resolves, the antilymphangiogenic effect increases. The balance between the pro-lymphangiogenic factors (VEGF-A, VEGF-C, VEGF-D) and anti-lymphangiogenic factors (Th1: IFN-g, Th2: IL-4, IL-13) produced by immune cells influences the actual response of lymphatic vessels to inflammatory stimuli. Accordingly, lymphatic vessels either grow or regress. In the clinic, treatments for lymphedema include lymphatic compression, exercise, or surgical interventions to improve the lymph flow. Thus, these research studies suggest that targeting the anti-lymphangiogenic factors alone or in conjunction with other therapeutic approaches could be a new strategy for treating lymphedema caused by infection or environmental stress.
Go to :
LYMPHATIC VESSELS IN MUCOSAL TISSUES
Lymphatics are prevalent in mucosal tissues that come into direct contact with the external environment. In this context, depending on the tissue environmental stimulation, LECs have distinct phenotypic and functional characteristics (30). Additionally, lymphatic vessels, which continuously transport circulating antigens, may represent an important line of defense against pathogens. It is striking that NF-κB activity is constitutive in lymphatics of respiratory and gastrointestinal systems that are in direct contact with external environment (31). In mice with a reporter gene for NF-κB activity (κB-lacZ), NF-κB is constitutively active in the lymphatic endothelium rather than in blood vessels in the uterus, intestine, and airways under normal conditions (31). These observations challenge the notion that the recognition of externally stimulated PAMPs by TLRs in lymphatic vessels results in the activation of transcription factor NF-κB as part of innate immunity. Several studies have revealed that LECs express large amounts of various TLRs and activate NF-κB by LPS and other TLR ligands for induction of inflammatory lymphangiogenesis (32). Therefore, LECs may have tissue-specific phenotypes and functions depending on the tissue environment in which they are located.
Gastrointestinal tract
Lymphatics in the gastrointestinal (GI) tract perform important mucosal immune functions in acquired immunity and tolerance because the GI tract is widely exposed to commensal bacteria. Lymphatic capillaries in the center of intestinal villi, called lacteal, are important for the absorption of nutrients as well as immune cell trafficking into mesenteric lymph nodes. The intestine contains three layers of lymphatics, including (in the direction from the luminal surface to the serosal surface): the lacteals in the villi, a submucosal lymphatic network, and a lymphatic network in the smooth muscle layer (Fig. 2). The lacteals connect with the submucosal lymphatic network at the base of the villi. A recent study revealed that lacteal longitudinally has been surrounded by bundles of smooth muscles and consequently are responsible for spontaneous contractile dynamics (33). This finding indicates that even though the lacteal is located at the end of peripheral tissue as a capillary, the lacteal has a unique tissue-specific lymphatic system that does not serve as a passive conduit but as an active pump, which transports lipids. Using conditional ablation of Lyve-1+ lacteals in the small intestine, it was revealed that the disappearance of lacteals gives rise to the distortion of blood capillaries and the whole architecture of villi, which allows for a massive invasion of intestinal pathogens without a barrier and immune surveillance and subsequently causes severe inflammation in the intestine (34). Likewise, intestinal lacteals contribute to the defense system against mucosal pathogens in the lumen.
 | Figure 2The small-intestine lymphatic system. The small intestine contains two independent lymphatic networks that drain into a common network of collecting lymphatics. The lacteals, draining individual villi, connect with a network of lymphatics within the mucosal layer to form one distinct network (red), while the lymphatics of the mucosal muscle layer form another network (blue). Injection of tracers into these networks reveals free flow within each network but no exchange between networks. Both networks drain into contractile collecting lymphatics (yellow) located at the mesenteric border and outside of the intestine.
|
Intestinal lymphatics are involved in a variety of disease processes such as congenital malformations resulting in pathological dilation of lymphatic vessels (lymphangiectasia) and inflammation with dynamic lymphatic growth and remodeling (35). Increased lymphatics have are seen in ulcerative colitis (UC) along with lymphangiectasia, particularly in areas with epithelial dysplasia (36). While lymphangiogenesis and lymphangiectasia are also found in ileal and colonic Crohn's disease (CD) (37), these observations imply a more direct correlation between lymphatics and the colitis pathogenesis. These studies showing a role of lymphatics in inflammatory bowel disease suggest that understanding the cause and consequences of lymphangitis could be a key to understanding Crohn's disease pathogenesis.
The intestinal environmental factors selectively affect the development of the intestinal lymphatic system. Fasting-induced adipose factor (Fiaf) also called angiopoietin- like protein 4 (Angptl4) is produced in the intestine and regulates the expression of Prox1, a transcription factor crucial for lymphangiogenesis (38). Deletion of the fiaf gene results in failure of blood and lymphatic vasculature partitioning in the small intestine. This finding emphasizes the uniqueness of the intestinal lymphatic system related to its role as a pathway for nutrient transport. Accumulating recent discoveries about the intestinal lymphatic vasculature in the GI tract offer convincing evidence that the lymphatic system in the intestinal mucosa is affected by and directly contributes to the pathogenesis of intestinal diseases.
Respiratory tract
In the steady state, lymphatic vessels of the respiratory tract optimize and facilitate the entrance of immune cells and interstitial fluid. As described above, the initial lymphatic capillaries are discontinuous button-like junctions, through which inflammatory cells frequently pass. Interestingly, the airway infected with Mycoplasma pulmonis shows growing lymphatic sprouts, which have a continuous junctional pattern, zippers instead of buttons (39). Such remodeling of the lymphatic vessels makes them less permeable and contributes to impaired fluid clearance and mucosal edema. After inflammation is resolved, the number of the lymphatic sprouts is decreased, and their microstructure again consists of buttons (40). Chronic inflammatory diseases involving the respiration system may be accompanied by significant structural alterations of lymphatic vessels, which probably are irreversible changes that are resistant to lymphatic-modulating treatments. When lymphangiogenesis is impaired, airway inflammation may lead to bronchial lymphedema and exaggerated airflow obstruction. Impaired drainage of the edema fluid may be expected to lead to airway wall thickening, reduced wall compliance, and narrowing of the airway lumen. Correction of defective lymphangiogenesis may benefit the treatment of asthma and other inflammatory airway diseases (41). Consistent with these consequences of impaired lymphangiogenesis is the finding that blocking of the lymphatic transport from inflamed rabbit ear skin may exacerbate local edema and eventually lead to necrosis (42). Thus, if lymphangiogenesis is impaired in inflamed airways, restoration of lymphangiogenesis may complement conventional therapies for inflammatory airway diseases.
Go to :
CONCLUSIONS
The lymphatic vasculature system, which has a specialized network of structures to transport tissue fluid and materials such as proteins and lipids to blood vasculature, closely interacts with distinct immune cells for execution of specific functions. For efficient flow from lymphatic capillaries to lymph nodes, LECs interact with myeloid cells in adipose tissue to regulate lymphatic permeability. For an immune reaction, activated immune cells such as DCs and memory T cells can actively or passively migrate from the tissue to lymph nodes through lymphatic vessels. The latter participate in the immune response by releasing an appropriate chemokine and cytokines in response to environmental changes in peripheral tissues. Because lymphatic vasculature is widespread throughout the whole body including the brain, it has been proposed that lymphatic vasculature maintains the peripheral tolerance in a steady state. In a pathological condition, however, inflammation frequently induces lymphangiogenesis, resolving inflammation by recruiting additional immune cells. Although VEGF-C and VEGFR3 have been mainly studied as key pro-lymphangiogenesis regulatory factors, it is still unknown whether other factors are involved in a tissue-dependent manner. In particular, intestinal lymphatic capillaries are constantly exposed to the commensal microbiota and are important for immunecell migration as well as nutrient absorbance. Therefore, it is necessary to conduct a molecular mechanistic study on the interaction between the commensal microbiota and lacteal development. Furthermore, the correlation between the lymphatic system and immune cells can be considered a therapeutic target in disease models such as Crohn's disease and ulcerative colitis in future studies.
Go to :
 XML Download
XML Download