

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
EPO
eosinophil peroxidase
SIRPα
signal regulatory protein α
ILC2 cells
type 2 innate lymphoid cells
LP
lamina propria
PPs
Peyer's patches
TFH cells
follicular B helper T cells
IL-1Ra
IL-1 receptor antagonist
BAFF
B cell activating factor of the tumor-necrosis factor family
APRIL
proliferation-inducing ligand
DCs
dendritic cells
Th1
IFN-γ-producing CD4+ T cells
Th2
IL-4-producing CD4+ T cells
Th17
IL-17-producing CD4+ T cells
Treg cells
regulatory CD4+ T cells
M1 macrophages
classically activated macrophages
M2 macrophages
alternatively activated macrophages
UCP-1
uncoupling protein-1
INTRODUCTION
Eosinophils are potent effector cells implicated in allergic responses and helminth infections and have cytotoxic granules containing major basic protein (MBP), eosinophil cationic protein (ECP), eosinophil peroxidase (EPO), and eosinophil-derived neurotoxin (EDN) (1). Responding to stimuli such as an allergen, helminth infection, or tissue injury, they release their granule-derived cytotoxic proteins and are involved in inflammatory processes. However, the intestine (constantly in contact with the gut microbiota) is a major site where eosinophils are abundantly present under homeostatic conditions. Intestinal eosinophils rarely undergo degranulation and have a long lifespan as compared with eosinophils in the blood and lungs (23). A possible reason is that inhibitory receptor signal regulatory protein α (SIRPα)/CD172a, highly expressed on the intestinal eosinophils, inhibits degranulation of eosinophils and promotes their survival (2). In support of this notion, the ligand for SIRPα/CD172a, CD47, is expressed in the intestine (2). In addition, intestinal eosinophils highly express common γ-chain, which is an integral part of the cytokine receptors for IL-2, IL-4, IL-7, IL-9, IL-15, and IL-21; intestinal eosinophils are severely reduced in number in the common γ-chain-deficient mice unlike eosinophils in blood and lungs; therefore, common γ-chain-dependent signals seem to play an important role in selective survival of intestinal eosinophils (3). Intestinal eosinophils are produced from hematopoietic stem cells in bone marrow and migrate into the intestine under the influence of chemokine eotaxin-1 (14). During the development and survival of eosinophils, signaling of the common β-receptor chain shared by IL-3, IL-5, and GM-CSF is required (4), and among these cytokines, IL-5 is particularly important; overproduction of IL-5 in transgenic mice results in eosinophilia (5) whereas deletion of IL-5 abrogates eosinophilia induced by an aeroallergen (6). According to a recent report, IL-5 is mainly produced by type 2 innate lymphoid (ILC2) cells, and its production in ILC2 cells is enhanced by vasoactive intestinal peptide (7). Additionally, transcription factor GATA-1 is crucial for the development of eosinophils; deletion of the palindromic double GATA high-affinity binding sites in the GATA-1 promoter results in eosinophil-deficient mice (called Δdbl GATA-1 mice) (8). PHIL mice, which express diphtheria toxin A chain under the control of the eosinophil-specific EPO promoter, are also eosinophil-deficient mice (9). In recent studies on these eosinophil-deficient mice, it has been revealed why eosinophils are abundant in the intestine without inflammation and what their physiological roles are under homeostatic conditions. Some papers have shown that intestinal eosinophils promote formation and maintenance of IgA-expressing plasma cells (10) and abundantly produce IL-1 receptor antagonist (IL-1Ra) (11), contributing to intestinal immune homeostasis. Several lines of evidence have shown that eosinophils also exist in adipose tissue and are involved in metabolic homeostasis (1213). Therefore, in this review, we summarize recent findings regarding eosinophils and discuss the homeostatic functions of these cells.
THE ANTI-INFLAMMATORY fUNCTION
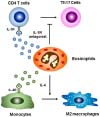
Recently, it has been reported that small-intestinal eosinophils produce large amounts of IL-1Ra (11). The latter competes with IL-1β for receptor binding and inhibits inflammatory responses (14). Meanwhile, IL-1β is primarily produced by activated macrophages and functions as a key mediator in various inflammatory diseases including inflammatory bowel disease and rheumatoid arthritis (14). In this context, a decrease in the IL-1Ra to IL-1 ratio is linked to inflammatory bowel disease in humans (15). IL-1Ra-deficient mice spontaneously develop arthritis with a marked increase in the number of Th17 cells (1617). In line with this finding, it has been reported that microbiota-induced IL-1β, but not IL-6, plays an important role in the development of Th17 cells in the intestine (18). Therefore, small-intestinal eosinophils suppress the differentiation and maintenance of Th17 cells by constantly secreting large amounts of IL-1Ra, thus contributing to intestinal immune homeostasis (11) (Fig. 1). Intestinal eosinophils also inhibit differentiation and/or proliferation of Th1 cells in the lamina propria (LP) though less strongly in comparison with the effects on Th17 cells (11). However, because IL-1Ra-deficient mice do not show a significant increase in the number of Th1 cells, intestinal eosinophils may suppress differentiation and/or proliferation of Th1 cells by secreting other unidentified cytokines. Unlike Th1 and Th17 cells, small-intestinal eosinophils do not affect frequencies of Foxp3+ regulatory T (Treg) cells and Th2 cells in the LP (11). Particularly in food allergy, intestinal eosinophils activate dendritic cells (DCs) through a release of EPO and promote their migration from the intestine to draining lymph nodes, thus initiating a primary Th2 immune response (19). In contradiction to these findings, one group reported that the numbers of CD103+ DCs and CD103+ CD4+ T cells including Foxp3+ Treg cells are reduced in the LP of eosinophil-deficient mice, although they did not explain the mechanism in the paper (10), and this phenomenon should be confirmed later.
Because small-intestinal eosinophils, but not eosinophils in blood and bone marrow, produce a large amount of IL-1Ra under homeostatic conditions, we can hypothesize that gut microbiota and its metabolites may be involved in IL-1Ra production. On the other hand, small-intestinal eosinophils in both SPF and GF mice produce similar levels of IL-1Ra, and metabolites such as retinoic acid and short-chain fatty acids do not induce IL-1Ra production in bone marrow eosinophils (11). Instead, GM-CSF, which is known to be involved in the development and survival of eosinophils, performs an important function in IL-1Ra production (11). In this regard, the high basal concentration of GM-CSF in the small intestine and high expression of GM-CSF receptor in small-intestinal eosinophils seem to explain the unique ability of small-intestinal eosinophils to continuously secrete a large amount of IL-1Ra under steady-state conditions. In addition, because Th17 cells can produce GM-CSF in an IL-1- and IL-23-dependent manner (20), it is plausible that GM-CSF produced by Th17 cells endows eosinophils with the superior capacity for IL-1Ra production and regulates Th17 cell homeostasis via a negative feedback loop. In contrast, ironically, large-intestinal eosinophils, when activated by GM-CSF produced by Th17 cells, secret EPO as well as proinflammatory cytokines TNF and IL-13, and aggravate colitis (21). These findings imply that small-intestinal eosinophils are different from large-intestinal ones. In support of this notion, large-intestinal eosinophils produce much smaller amounts of IL-1Ra and show a distinctive expression pattern of surface proteins including CD11c, ST2, and Ly6C as compared with small-intestinal ones (19). In addition, although eosinophils are mainly present throughout the intestine, large-intestinal eosinophils are scarce in a steady state (1119), and only under intestinal inflammatory conditions, does their frequency dramatically increase (21). Accordingly, large-intestinal eosinophils seem to be proinflammatory unlike their small-intestinal counterparts.
ANTIBACTERIAL DEFENSE
Most of the intestinal plasma cells are located in the LP and mostly produce IgA (22). IgA is secreted into the intestinal lumen and functions as the first line of defense through neutralization and clearance of enteric pathogens. Nevertheless, IgA-producing B cells are generally formed in Peyer's patches (PPs) and migrate into the LP (22). In PPs, antigen-experienced CD4+ T cells migrate into B-cell follicles via CXCR5 and activate B cells (22). These specialized CD4+ T cells are called follicular B helper T (TFH) cells and are different from other CD4+ T cell lineages. They highly express costimulatory molecules for their development and function, e.g., CD40L, OX-40, and programmed cell death 1 (PD-1) rather than cytokines and transcription factors characteristic of Th1, Th2, and Th17 cells (22). Transcription factor B cell lymphoma 6 (Bcl-6) plays an important role in TFH differentiation, whereas transcription factor B lymphocyte maturation protein 1 (Blimp-1) inhibits it (22). The activated B cells induce activation-induced cytidine deaminase (AID) and provoke class switch recombination (22). Because TGF-β1, a key cytokine for IgA switching, is expressed by many cells in PPs, activated B cells preferentially switch their isotype from IgM to IgA (2223). Moreover, IL-21 secreted by TFH cells enhances the proliferation and differentiation of IgA plasma cell precursors in synergy with TGF-β1 (24). Meanwhile, IgM+ B cells in the LP are activated by polyclonal stimuli or by antigens presented by DCs in the LP (LP-DCs) and differentiate into IgA-producing plasma cells without the help of TFH cells (22). In this T cell-independent formation of IgA-producing B cells, important functions are performed by TGF-β1, B-cell activating factor of the tumor-necrosis factor family (BAFF), and a proliferation-inducing ligand (APRIL) produced by DCs and stromal cells in the LP (22). In vitro, cytokines such as IL-6, TNF, IL-10, IL-4, and IL-5, also prolong the survival of plasma cells (25). In this context, bone marrow eosinophils have been reported to support the survival of plasma cells by secreting APRIL and IL-6 and contribute to the long-term maintenance of plasma cells in bone marrow (26). Although intestinal eosinophils are mainly in the LP, not in PPs, they are involved in the formation and maintenance of IgA-producing B cells in PPs as well as in the LP (10). Intestinal eosinophils express BAFF, TGF-β1, and matrix metalloproteases (that are involved in the release of TGF-β1 from the large latent complex) as well as APRIL and IL-6 at the mRNA level (10). Therefore, intestinal eosinophils appear to influence T-cell-independent formation and maintenance of IgA-producing B cells via production of these cytokines and enzymes. Accordingly, the amount of IgA is dramatically reduced in the intestine of eosinophil-deficient mice; this change increases bacterial load in the gut and affects composition of the microbiota there (10). Therefore, it seems likely that eosinophils indirectly exert their effects on T-cell-dependent formation of IgA-producing B cells in PPs. Recently, one paper revealed that eosinophils can be involved in IgA production through IL-1β (27). Moreover, eosinophils can release mitochondrial DNA in a catapult-like manner and kill bacteria (28). Nevertheless, the release of mitochondrial DNA does not cause cell death of eosinophils. For the release of mitochondrial DNA, eosinophils should be activated by lipopolysaccharide under conditions where they are primed with IL-5 or IFN-γ (28). In the intestinal environment, gut bacteria can exert their effects on immune cells through bacteria-derived molecules, and immune-cell populations producing IFN-γ (Th1) and IL-5 (ILC2) exist; therefore, it seems plausible that intestinal eosinophils reduce the bacterial load in the gut via the release of mitochondrial DNA under homeostatic conditions. Accordingly, intestinal eosinophils prevent the host from bacterial invasion and contribute to homeostasis of intestinal immunity.
METABOLIC HOMEOSTASIS
Obesity induces chronic low-grade inflammation, consequently causing insulin resistance and type 2 diabetes (2930). In the obese state, a large number of monocytes migrate into adipose tissue and differentiate into classically activated (M1) macrophages expressing TNF-α and inducible nitric oxide synthase, which triggers inflammation and results in metabolic diseases (2930). By contrast, resident adipose-tissue macrophages in the lean state express IL-10 and arginase 1, and show an anti-inflammatory phenotype of alternatively activated (M2) macrophages (2930). Activation of the anti-inflammatory M2 macrophages requires Th2 cytokines such as IL-4 and IL-13 (Fig. 1), whereas activation of M1 macrophages requires Th1 cytokine IFN-γ (2930). Eosinophils are a major IL-4-expressing cell population in adipose tissue and help to sustain M2 macrophages (12) (Fig. 1). High-fat diet-induced obesity significantly reduces the number of eosinophils in adipose tissue (12). Therefore, eosinophil-deficient mice show aggravation of obesity-induced metabolic diseases via a dramatic reduction in the number of M2 macrophages in adipose tissue, indicating that eosinophils are intimately involved in metabolic homeostasis (12). Besides, ILC2 cells promote accumulation of eosinophils and activate M2 macrophages in adipose tissue through production of IL-5 and IL-13 and are implicated in metabolic homeostasis (3132).
Eosinophils are also involved in the biogenesis of beige fat (1333). The latter represents clusters of uncoupling protein 1 (UCP-1)-expressing adipocytes in white adipose tissue and functions in heat production under conditions of increased energy expenditure, such as cooling and exercise (3435). Thus, beige adipocytes contribute to suppression of obesity and alleviation of metabolic diseases. By contrast, white adipocytes do not express UCP-1 and are not involved in thermogenesis; instead, they participate in fat storage, contributing to obesity-induced metabolic diseases (35). IL-4 released by eosinophils induces the tyrosine hydroxylase expression of M2 macrophages under cold conditions and drives these macrophages to produce catecholamines such as norepinephrine (13). The macrophage-derived catecholamines trigger expression of the UCP-1 gene in subcutaneous white adipose tissue and are involved in the development of beige fat (13). In addition, meteorinlike (Metrnl), which is a hormone induced in muscle after exercise and in adipose tissue upon cold exposure, exerts its effect on eosinophils and increases their expression levels of Th2 cytokines such as IL-4 and IL-13, consequently promoting the development of (and thermogenesis in) beige fat (36). In particular, ILC2 cells contribute to the biogenesis of beige fat through production of methionine-enkephalin peptide as well as via activation by IL-33 receptor signaling (32).
Increasing evidence suggests that obesity and other metabolic syndrome are associated with altered composition of the gut microbiota (37). In agreement with this notion, the transfer of gut microbiota from obese to germ-free mice increases obesity more than gut microbiota from lean mice does (38). Microbiota depletion either by antibiotic treatment or in germ-free mice promotes beige fat biogenesis and alleviates obesity and other metabolic syndrome; these effects are mediated by eosinophil accumulation, enhanced Th2 cytokine signaling, and M2 macrophage polarization in white adipose tissue of microbiota-depleted mice (39). In this regard, intestinal eosinophils may inhibit obesity and other metabolic syndrome through a reduction of bacterial burden via IgA production (1027). In addition, because obesity-induced metabolic diseases show elevated levels of IL-1β (29), intestinal eosinophils may alleviate inflammation by producing large amounts of IL-1Ra and thus alleviate metabolic diseases.
PROSPECTS
According to recent reports, eosinophils are a heterogeneous cell population and have different characteristics depending on the site of residence (3111921). Although eosinophils are mainly distributed in the intestine, the phenotype of large-intestinal eosinophils is different from that of small-intestinal ones, and the number of large-intestinal eosinophils in a steady state is much smaller than that of their small-intestinal counterparts (111921). Moreover, small-intestinal eosinophils in response to GM-CSF produce large amounts of IL-1Ra and exert an anti-inflammatory function (11), whereas large-intestinal eosinophils increase production of proinflammatory cytokines TNF-α and IL-13 and promote colitis (21). These observations mean that eosinophils do not all have an identical function, but rather their function is likely controlled by local milieu. Thus, if anti-inflammatory eosinophils can be upregulated or inflammatory eosinophils can be converted into anti-inflammatory ones, then these modalities should help to combat inflammatory diseases such as allergies, inflammatory bowel disease, and obesity-related metabolic diseases. To this end, it would be worthwhile to identify the surface markers that can distinguish homeostatic and inflammatory eosinophils. The extract of Ulmus davidiana var. japonica Nakai increases the number of small-intestinal eosinophils and suppresses differentiation and/or proliferation of Th1 and Th17 cells (40). Regulatory eosinophil-recruiting prebiotics or probiotics may be useful for prevention of inflammatory and metabolic diseases. More studies should be conducted in this field for successful development of eosinophil-targeting treatments.
 XML Download
XML Download