

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
DC
dendritic cell
iBALT
induced bronchus-associated lymphoid tissue
LTα
lymphotoxin α
LTβ
lymphotoxin β
RORγt
retinoic acidrelated orphan receptor gamma t
LTi
lymphoid tissue inducer
TH17
T helper 17
S1P
sphingosine-1-phosphate
PAO
protease from Aspergillus oryzae
BMDC
bone marrow-derived dendritic cell
mLN
mediastinal lymph node
INTRODUCTION
T cell clonal expansion is an explosive increase of antigen-specific T cells which are essential for efficient adaptive immunity against invading foreign antigens (1,2). The clonal expansion initiates when antigen-specific naive T cells engage dendritic cells (DCs) bearing a cognate antigen in the secondary lymphoid organs such as lymph nodes (3,4). The recognition of an antigenic peptide in the context of MHC molecules by a T cell receptor delivers antigen-driven signals into T cells, termed ‘signal 1’. Costimulatory molecules on the antigen-presenting DCs provide additional signals (signal 2) that lead to clonal expansion by inducing autocrine IL-2 production from T cells. In addition, cytokines from the DCs further stimulate the activated T cells to differentiate into a certain lineage of effector T cells (signal 3) (567).
It is well-established that the clonal expansion of naïve T cells occurs in the draining lymph nodes since antigen-bearing DCs migrate into the T cell zone of the lymph nodes by upregulating C-C chemokine receptor type 7 (8,9). On the other hand, T cell-DC interaction can also occur in the tertiary lymphoid structures in non-lymphoid organs such as the lung. For instance, induced bronchus-associated lymphoid tissue (iBALT) is one type of tertiary lymphoid structures in the lung. Recent studies highlighted a crucial role of this tertiary lymphoid structure in the priming of T cell responses during influenza A or herpesvirus infection (10,11). However, whether inhaled allergens can also trigger the clonal expansion of naïve T cells in the lung remains unknown.
Lymph node-deficient mice are useful tools in studying the requirement of the lymph node in inducing immune responses. Mice deficient in lymphotoxin α (LTα) exhibit abnormal development of peripheral lymphoid organs. These mice do not have detectable popliteal, inguinal, para-aortic, mesenteric, axillary, or cervical lymph nodes (12). LTα-deficient mice show normal counts of CD4+ and CD8+ T cells in peripheral blood. On the other hand, Lymphotoxin β (LTβ) -deficient mice have mesenteric and cervical lymph nodes (13). Tumor necrosis factor (TNF) is a member of TNF ligand family and is structurally similar to LTα and LTβ. Lta/Ltb/Tnf triple knock-out mice also have abnormal lymphoid development similar to that of the phenotype of LTα- or LTβ-deficient mice. Unlike single mutant mice, these mice have additional disruption to the spleen microarchitecture and exhibit impaired antibody responses to a T cell-dependent antigen (14). The retinoid acid-related orphan receptor gamma t (RORγt) is required for the development of lymphoid tissue inducer (LTi) cells and for the generation of T helper 17 (TH17) cells (15). The Rorgtgfp/gfp mice are defective of all lymph nodes and Peyer's patches. The mice are protected from the experimental autoimmune encephalomyelitis due to the lack of tissue-infiltrating TH17 cells (16,17).
In this study, we aimed to investigate whether the clonal expansion of allergen-specific T cells can occur in the lung by using lymph node-deficient Rorgtgfp/gfp mice and sphingosine-1-phosphate (S1P) antagonist FTY720 in an intranasal allergen-induced animal model of lung inflammation in vivo. Our findings demonstrate that the clonal expansion of naïve CD4+ T cells can occur in the lung in response to the exposure of inhaled allergens.
MATERIALS AND METHODS
Mice
C57BL/6 mice were purchased from Orient. Rorgtgfp/gfp , OT-II and B6.SJL (CD45.1+) mice were purchased from Jackson Laboratory. OT-II mice were crossed with B6.SJL for adoptive transfer study. All mice were maintained under the semi-specific-pathogen-free facility in an animal center (Seoul National University). All animal experiments were performed using a protocol approved by the Institutional Animal Care and Use Committee Seoul National University (SNU-140602-2-10).
Allergen-induced lung inflammation
Mice were anesthetized with isoflurane and were intranasally administered with a mixture of 7 µg of proteinase from Aspergillus oryzae and 20 µg of ovalbumin (PAO/Ova) in 50 µL PBS every other day for a total of four times (day 0, 2, 4, 6). Sixteen hours after the last challenge, all mice were euthanized and the mediastinal lymph node, the lung and blood were collected for further analysis.
CFSE-labeled OT-II adoptive transfer
OVA-specific CD4 T cells were isolated from the lymph nodes and the spleens of B6.SJL OT-II mice by using a CD4+ T cell Isolation Kit. The isolated CD4+ T cells were labeled with 2 µM of CFSE and intravenously transferred into C57BL/6 mice. Next day, the recipients were intranasally injected with PAO/Ova (50 µg/7 µg). For kinetic analysis experiments, the mediastinal lymph node and the lung were dissected from mice after 36, 48, 60, 72 hours from challenge and the CFSE+ cells were analyzed using flow cytometry.
FTY720 treatment
For T cell migration inhibition experiments, CFSE-labeled OT-II cells (5×106 cells/transfer) were transferred into C57BL/6 mice and then the mice were treated with intraperitoneal injection of FTY720 (1 mg/kg) or vehicle 6 hours after intranasal administration. Forty-eight hours after the intranasal administration, all mice were euthanized and the mediastinal lymph node, the lung and blood were obtained and analyzed by flow cytometry.
Isolation of lymphocytes from the lung and blood
For lymphocyte isolation from mouse lung, the lung was dissected into single lobes and cut into small pieces using the Gentle MACS Dissociator. These lobes were digested in RPMI 1640 medium containing 10% FBS, 0.5 mg/mL of collagenase IV, 2 mg/mL of dispase, and 2.5 µg/mL of DNase I for 30 minutes at 37℃. The lung cells were filtered (100 µm) and then washed with PBS containing 1.5% FBS. The lymphocytes were isolated from whole lung cells or heparinized blood using Lymphocyte Separation Medium.
Flow cytometry
For the flow cytometry analysis, cells were stained with PE-Cyanine7-conjugated anti-mouse CD4 (RM4-5), Per-CP/Cy5.5-conjugated anti-mouse CD45.2 (104), Pacific Blue-conjugated anti-mouse CD45.1 (A20) and APC-conjugated anti-mouse TCR Vα2 (B20.1). These cells were analyzed by FACSVerse and obtained data were analyzed using a software called Flowjo.
CD4+ T cell proliferation in vitro
CD4+ cells were isolated from lymph nodes and spleen of wild-type and Rorgtgfp/gfp mice with CD4+ T cell Isolation Kit and then CD4+CD62L+CD25–CD44– cells were sorted using FACSAria III cell sorter. These isolated naïve T cells were labeled with 2 µM of CFSE. The CFSE-labeled naïve T cells (1×105/well) were co-cultured with bone marrow-derived dendritic cells (BMDC) (1×104/well) in RPMI-1640 supplemented with 10% FBS and soluble anti-CD3 (2C11, 0.5 µg/mL) for 72 hours.
RESULTS
Analysis of CD4+ T cells in the lung of lymph node-deficient Rorgtgfp/gfp mice challenged with intranasal allergens
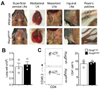
Draining lymph nodes harboring antigen-captured DCs act as primary sites of the proliferation of T cells specific to the antigen, termed clonal expansion. As a first step to determine if lymph nodes are necessary to initiate allergen-specific CD4+ T cell responses in the airway, we employed Rorgtgfp/gfp mice in which all peripheral lymph nodes and Peyer's patches are absent due to the lack of LTi cells (Fig. 1A) (15). Intranasal challenges with proteinase from Aspergillus oryzae in combination with ovalbumin (PAO/Ova) have been used to induce allergic airway inflammation in mice (181920). Total number of immune cells as well as the frequency of CD4+ T cells in the lung of unchallenged mice was comparable between Rorgt+/gfp and Rorgtgfp/gfp mice (Fig. 1B and 1C). We intranasally challenged wild-type or Rorgtgfp/gfp mice with PAO/Ova every other day for a total of four times (Fig. 2A). One day after the last challenge, we analyzed immune cell population in the airway and found that the majority of CD45.2+ cells in the lung of Rorgtgfp/gfp mice were CD4+ T cells. Although the result was not statistically significant, the frequency of CD4+ T cells in the lung of Rorgtgfp/gfp mice was slightly lower in comparison to that of wild-type mice (Fig. 2B). These results suggest that the CD4+ T cell responses against allergen challenges could occur in the airway even in the absence of lymph nodes.
Kinetic analysis of allergen-specific CD4+ T cell division in the BALT upon intranasal allergen
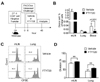
To characterize the expansion of CD4+ T cells in the bronchus-associated lymphoid tissues (BALT) against intranasally-challenged allergens, we performed kinetic analysis of allergen-specific T cell proliferation to intranasal allergen. We adoptively transferred CFSE-labeled CD4+ T cells from B6.SJL OT-II mice (CD45.1+CD45.2+) into the C57BL/6 recipients (CD45.2+) before the recipients were challenged with a single dose of intranasal PAO/Ova. The dilution of CFSE in the donor T cells in the mediastinal lymph node (mLN) and the lung of the recipients was analyzed in 36, 48, 60 and 72 hours after the intranasal challenge (Fig. 3A). As shown in Fig. 3B, after 36 hours the evident division of OT-II T cells was observed in the mLN, but not in the lung. By 48 hours, the majority of donor OT-II T cells in the mLN as well as in the lung underwent at least one division (Fig. 3C and 3D). Interestingly, we observed no difference in the percentage of donor T cells that divided more than once between the mLN and the lung at 48 hours and thereafter. These data suggest that the expansion of allergen-specific CD4+ T cells occurred primarily in the mLN, but the division number of donor cells in the mLN and in the lung was appeared to be comparable by 48 hours after allergen challenge.
Expansion of allergen-specific CD4+ T cells in the lung of FTY720-treated mice
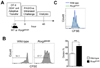
The observation in Fig. 2 and Fig. 3 suggested that the clonal expansion of allergen-specific CD4+ T cells might occur in the lung in the absence of lymph nodes. To explore this hypothesis, we utilize an antagonist of S1P receptor, FTY720, which blocks the S1P-dependent egression of lymphocytes from the lymph nodes. FTY720 does not affect the proliferation of antigen-specific T cells (21,22). CFSE-labeled OT-II T cells (CD45.1+CD45.2+) were intravenously injected into C57BL/6 recipients and the recipients were intranasally challenged with PAO/Ova. Since FTY720 can also affect the migration of DCs (232425), we injected FTY720 or vehicle into the recipients 6 hours after the intranasal challenge. Forty-eight hours after the challenge, we analyzed the dilution of the CFSE in the donor T cells (Fig. 4A). As depicted in Fig. 4B, the percentage of donor T cells in the mLN and the lung was found to be comparable between vehicle-and FTY720-treated groups. However, the frequency of donor T cells in the blood was profoundly lower in the FTY720-treated mice compared to that of vehicle-treated mice, indicating that such treatment efficiently inhibited the egression of allergen-specific T cells from the mLN.
Notably, we observed an evident donor OT-II T cell population that underwent at least one division in the lung of FTY720-treated recipients, and that the percentage of these dividing donor OT-II T cells was slightly, but significantly, higher in the FTY720-treated recipients than that of vehicle-treated recipients (Fig. 4C and 4D). Similar results were observed in the mLN of the recipients. These results demonstrate that the clonal expansion of allergen-specific CD4+ T cells in response to intranasal allergens can occur simultaneously in the lung and in the mLN in mice treated with FTY720.
Expansion of allergen-specific CD4+ T cells in the lung of Rorgtgfp/gfp mice
The observed division of OT-II T cells in the lung of FTY720-treated mice in Fig. 4 prompted us to hypothesize that the clonal expansion of allergen-specific CD4+ T cells could be initiated in the absence of egression of activated T cells from the mLN in vivo. To explore this possibility in a definitive in vivo system, we asked if the expansion of allergen-specific CD4+ T cells in the lung could occur in Rorgtgfp/gfp mice lacking lymph nodes. We performed a similar adoptive transfer study with CFSE-labeled OT-II T cells (CD45.1+CD45.2+) into wild-type or Rorgtgfp/gfp mice, and the recipients were intranasally challenged with PAO/Ova (Fig. 5A).
Consistent with the observation in Fig. 4, we observed an evident division of the donor OT-II T cells in the lung of Rorgtgfp/gfp mice (Fig. 5B). Unexpectedly, the frequency of donor T cells that underwent at least one division in the lung of Rorgtgfp/gfp mice was significantly higher than that of wild-type mice (Fig. 5B). We observed no difference in the proliferation of wild-type and Rorgtgfp/gfp CD4+ T cells in vitro (Fig. 5C). This indicates that the division of OT-II T cells in the lung was accelerated in the Rorgtgfp/gfp mice, probably due to the lack of mLN. Collectively, these results imply that the clonal expansion of allergen-specific CD4+ T cells occurred in the lung independently of the lymph nodes in vivo.
DISCUSSION
DCs residing in the lung capture inhaled allergens and migrate into the draining lymph nodes. The allergen-specific adaptive immune responses are initiated by the interaction between these allergen-bearing DCs and allergen-specific CD4+T cells in the lymph nodes, which results in an expansion of the allergen-specific T cell clones. However, whether lung can act as a primary site for the clonal expansion of allergen-specific CD4+ T cell has been unclear. Our findings in the present study collectively indicate that the clonal expansion of allergen-specific CD4+ T cells could occur in the lung in the absence of the lymph nodes, since the division of allergen-specific CD4+ T cell takes place in the lung of mice treated with an S1P receptor antagonist FTY720 as well as in the lung of Rorgtgfp/gfp mice lacking all peripheral lymph nodes. Hence, although the draining lymph nodes are the primary sites of clonal expansion of T cells under steady-state, allergen-specific clonal expansion can also occur in the absence of the lymph nodes in vivo.
Immature DCs residing in the lung capture invading antigens in the airway and migrate into the draining lymph nodes through chemotaxis where they differentiate antigen-specific naïve T cells into effector T cells by providing signal 1, 2 and 3. Effector T cells then leave the lymph nodes and migrate into the periphery to mediate adaptive immunity (26). Aside from the lymph nodes, tertiary lymphoid structures can be developed in the lung in case of chronic inflammation, infection and cancer. These tertiary lymphoid structures are ectopic lymphoid-like structures that consist of T cells and B cells (27,28). Like the lymph nodes, antigen-specific T cell responses are known to be induced in these tertiary lymphoid structures (27). In this regard, it is noteworthy that secondary lymphoid organs are not essential for the maintenance of immunologic memory and the utility of iBALT in a murine model of herpesvirus MHV-68 infection (11). In line with this study, our results in the present study showed that the expansion of allergen-specific T cell occurs in the lung in the absence of lymph nodes in Rorgtgfp/gfp mice and in the lung of FTY720-treated mice after intranasal challenges with allergens. It has been suggested that CD11c+ DCs are essential for the efficient priming of T cell responses against influenza A infection in the tertiary lymphoid structure. By contrast, it has been also shown that CD11c+ DCs are not necessary for the maintenance of tertiary lymphoid structures in an allergic airway inflammation model (10,11). The type(s) of antigen presenting cells that trigger clonal expansion of naïve CD4+ T cells in the lung in the present study is unclear at the moment.
Treatment with FTY720 has been shown to suppress the T helper 1- and T helper 2-driven lung inflammation by reducing migration of T cells from the mLN into the lung and by reducing the migration of lung DCs to the mLN (29,30). Although these prior findings seem to contradict the observed expansion of allergen-specific CD4+ T cells in the lung of FTY720-treated mice and Rorgtgfp/gfp mice, the question of whether the clonal expansion of allergen-specific CD4+ T cells can occur in the lung in the absence of the lymph nodes has not been addressed. Similar to our findings, it has been also shown that RORγt-deficient mice harbored an increased number of iBALT in response to influenza A virus infections (31). Taken together, these prior studies and the present study suggest that, when the lymph nodes are absent, allergen-bearing DCs in the lung could trigger T cell clonal expansion in the lung, probably by interacting circulating T cells. Further studies are necessary to dissect the exact cellular and spatial mechanisms of the interaction between DCs and naïve T cells in the lung.
In summary, the present study unveils that the clonal expansion of CD4+ T cells occurs in the lung in response to intranasal allergens in the absence of the lymph nodes in vivo. The contribution of this lymph node-independent expansion of allergen-specific T cells on the development of allergic airway inflammation needs to be determined.


 XML Download
XML Download