

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
α1-antitrypsin (AAT) exhibits anti-inflammatory activity and its deficiency causes respiratory and inflammatory disorders (12345678). AAT directly blocks serine proteinases activating interleukin (IL)-1 cytokine family, IL-8, IL-32, and tumor necrosis factor (TNF)-α (91011). IL-32, originally discovered as a natural killer cell transcript 4 (NK4) and renamed as IL-32 (also known as TNFα-inducing factor (TAIF) in databank), induces various inflammatory cytokines via NF-κB and p38MAPK signaling (1012131415161718). IL-32α-ligand affinity chromatography lead to isolation of neutrophil proteinase 3 (PR3) from human urine (9). Mature PR3 specifically binds to IL-32 and cleaves it. The suppression of IL-32 expression by AAT increased survival in an allogeneic bone marrow (BM) transplantation mouse model (19). Unlike other cytokines possessing specific membrane receptors, IL-32 utilizes membrane PR3 to induce inflammation. Therefore, this study aimed to investigate the inhibitory activity of AAT on IL-32-induced inflammation in mouse BM cells.
MATERIALS AND METHODS
Primary mouse BM cells were isolated from 6-week-old female C57BL/6 mice as described previously (20), seeded on a 96-well plate at a density of 1×106/0.2 mL, and then stimulated with 100 ng/mL LPS (Sigma-Aldrich, St. Louis, MO, USA), TNFα, IL-32γ, and IL-1α (YbdYbiotech, Seoul Korea) as indicated at the bottom. The animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of Konkuk University. IL-32γ-induced NF-κB and p38MAPK phosphorylation in BM cells isolated from 6-week-old C57BL/6 mice was examined using anti-phospho antibodies (Cell Signaling Technology, Beverly, MA, USA). Recombinant PR3-Fc was expressed as described previously (21), and then was used in IL-32γ binding assay. A 96-well plate was coated with PR3-Fc (1 µg/mL) overnight and blocked with 3% BSA for 2 h. IL-32γ was added at the concentration indicated at the bottom, and was detected by rabbit anti-IL-32γ antibody (22). AAT-Fc was pre-incubated with PR3-Fc-coated 96-well plate at a concentration indicated at the bottom, and then its reaction with IL-32γ was detected by rabbit anti-IL-32γ antibody.
RESULTS AND DISCUSSION
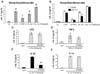
Here, the anti-inflammatory activity of AAT was investigated in BM cells. Primary mouse BM cells were stimulated with LPS, TNFα, IL-32γ, and IL-1α because they trigger common inflammatory pathways, such as NF-κB and p38MAPK, although they possess a specific receptor. IL-1α induced IL-6 at the highest level, whereas TNFα induced IL-6 at the lowest level, as compared to the induction level of other stimuli (Fig. 1A). Mouse BM cells were pre-incubated with recombinant AAT-Fc and then triggered with the same stimuli (Fig. 1B). Interestingly, AAT-Fc specifically suppressed IL-32γ-induced IL-6 expression, while AAT-Fc failed to inhibit IL-6 expression induced by other stimuli (Fig. 1B). This data is presented for the first time with direct evidence that AAT-Fc inhibits the inflammatory activity of IL-32 in mouse BM cells. In addition, AAT-Fc experiments were performed by pretreating mice for 48 h prior to isolating BM cells. As shown in Fig. 1E, IL-32γ-induced IL-6 expression was further decreased in AAT-Fc-pretreated mice, but IL-6 expression induced by other stimuli was not inhibited (Fig. 1C, D, and F).
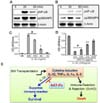
Next, NF-κB and p38MAPK signaling were assessed in the mouse BM cells after IL-32γ stimulation. The phosphorylation of NF-κB and p38MAPK was increased by IL-32γ treatment (Fig. 2A), while it was significantly suppressed in the BM cells from AAT-Fc-administered mice (Fig. 2B). In addition, recombinant PR3-Fc was expressed in Chinese Hamster Ovary cells by using the open reading frame of PR3 fused to human Fc as described previously (21). The PR3-Fc protein was used to indicate IL-32γ binding to PR3. IL-32γ binds to PR3-Fc in 1 nM range with a high affinity, since approximately 30 kDa IL-32γ exhibits a specific binding at 30 ng/mL (Fig. 2C). The specific IL-32γ binding to PR3-Fc was abrogated by AAT-Fc in a dose-dependent manner (Fig. 2D).
These data suggest that AAT-Fc blocks IL-32-induced inflammation in BM cells by competing with IL-32γ to bind on membrane PR3 and increases the survival of allogeneic BM transplantation mouse model (Fig. 2E). Unlike other cytokines with specific membrane receptors, IL-32 utilizes membrane PR3 to induce inflammation. This may explain the specific inhibitory activity of AAT on IL-32-induced inflammation in patients with allogeneic BM transplantation. The present study illustrates the function of IL-32 in BM inflammation that is specifically suppressed by AAT-Fc. Further investigation on IL-32-mediated inflammation in human BM transplantation will help understand the causative factor in GvHD.
 XML Download
XML Download