

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
AML
acute myeloid leukemia
ALL
acute lymphoblastic leukemia
AOM
azoxymethane
CRC
colorectal carcinoma
DUSP
dual-specificity phosphatase
DSS
dextran sodium sulphate
EGFR
epidermal growth factor receptor
ERK1/2
extracellular signal-regulated
protein kinase
JNK
c-Jun NH2-terminal kinase
LOH
loss of heterozygosity
MAPK
mitogen-activated protein kinases
MAPKK
MAPK kinase
miR
microRNA
NPC
nasopharyngeal carcinoma
SCC
squamous cell carcinoma
OVERVIEW OF THE MAPKs
The evolutionarily conserved mitogen-activated protein kinase (MAPK) cascades coordinately regulate a wide array of cellular processes such as cell proliferation, differentiation, metabolism, motility, survival, apoptosis, and are therefore important for numerous physiological processes including innate and adaptive immune responses (123).
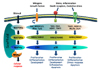
There are 3 main groups of MAPKs in mammalian species; the extracellular signal-regulated protein kinases (ERK1/2), the p38 MAP kinases (isoforms α, β, γ, δ) and the c-Jun NH2-terminal kinases (JNK1/2/3) (Fig. 1) (4). These MAPK pathways can be activated by various cell stimuli, including growth factors, cytokines and cellular stress, which leads to a cascade of sequential phosphorylation events, beginning with the phosphorylation of MAPK kinases (MAPKKs) on their Ser/Thr residues by MAPK kinase kinases (MAPKKKs) (5). Activated MAPKKs in turn phosphorylate MAPKs at the adjacent threonine and tyrosine residues in the conserved Thr-X-Tyr (TXY) motif within their activation loop. The amino acid denoted X corresponds to glutamic acid in ERK, proline in JNK or glycine in p38 (678).
MAPKs respond to a wide range of extracellular and intracellular changes. Activated MAPKs can phosphorylate/activate a wide array of downstream targets, including protein kinases and transcription factors that facilitate the transcription of MAPK-regulated genes (34910). The cellular response of the MAPK pathways depends on the nature of the stimulus and the duration of the signal (11). In general, the ERK pathway is mainly involved in growth, differentiation, and development, while JNK and p38 MAPK pathways are preferentially involved in inflammation, apoptosis, growth, and differentiation (10). In immune cells, the MAPK pathways are downstream targets of cell receptor signaling, working cooperatively to regulate various cellular responses such as cell proliferation, differentiation, effector function, and cell death (Fig. 1) (312).
MAPK SIGNALING PATHWAYS IN CANCER
Perturbations in MAPK signalling are a feature in cancer development and progression. Although the ERK1/2 pathway is commonly linked with cell growth, recent studies have implicated this pathway in other aspects of the phenotype of tumors. As such, aberrant ERK1/2 activation has been implicated in one-third of all human cancers (13). Most oncogenic activities that lead to constitutive activation of ERK1/2 signalling occur upstream of ERK1/2 activation, including overexpression of receptor tyrosine kinases such as epidermal growth factor receptor (EGFR), activating mutations in receptor tyrosine kinases, sustained autocrine or paracrine production of activating ligands, as well as Ras mutations and B-Raf mutations, resulting in enhanced or constitutive downstream activation of the Raf-MEK-ERK pathway (9131415). Phosphorylation of ERK1/2 via the Ras/Mek/ERK pathway cascade induces the activation of transcription factors NF-κB, AP-1, and ETS, resulting in the induction of downstream components, c-Fos, cyclin D1, and c-Myc, which are important cell proliferation and growth regulation factors (1617). Dysregulation of this pathway has been demonstrated in a variety of malignancies, including hepatocellular carcinoma, gastric adenocarcinoma, and renal cell carcinoma. Additionally, ERK1/2 promotes cell survival by inhibiting apoptosis in response to a wide range of stimuli, such as TNFα, Fas ligand, TRAIL, radiation, osmotic stress, hypoxia, growth factor withdrawal, nitric oxide, hydrogen peroxide, matrix detachment and chemotherapeutic agents (18).
The role of the stress activated MAPKs, including JNK and p38, in cancer is complex and in some cases controversial. Increased levels of phosphorylated p38 have been linked to various malignancies, including follicular lymphoma, lung, thyroid and breast carcinomas, as well as glioma and head and neck squamous cell carcinomas (2). In contrast, studies using mice with disrupted genes of the p38 kinases, MKK3 and MKK6, or the p38α gene demonstrated enhanced transforming potential of fibroblasts, indicating a role of the p38 MAPK pathway in tumor suppression (192021). The p38 pathway has also been implicated in the activation of p53 and p53-mediated apoptosis (22). Genetic inactivation of WIP1, a phosphatase stimulated by p53 and that can target p38, reduces mammary gland tumorigenesis in mice (20). This correlates with increased levels of p38 activity and apoptosis. In addition, p38 deficient mice are sensitized to KRAS-induced lung tumorigenesis, which was characterized by immature hyperproliferation of lung epithelial cells resulting from p38 inactivation (23).
The tumor suppressive functions of JNK are often related to their pro-apoptotic activity while the oncogenic functions of JNK depend on their ability to phosphorylate downstream target c-Jun and to activate the transcription factor, AP-1. Studies have indicated that JNK activity was required for Ras-induced cell transformation in vitro (24). In hepatocellular carcinoma (HCC), increased JNK1 activity has been associated with tumor cell proliferation and altered histone H3 methylation (2526). Similarly, JNK pathway activation and c-Jun phosphorylation have been associated with Ras-induced tumor development and cellular transformation (27). Ras and JNK have been found to phosphorylate c-Jun at the same sites and c-Jun-deficient fibroblasts have been reported to be resistant to Ras-dependent transformation (2829). In a mouse model of HCC, JNK1, but not JNK2, deficiency has been shown to reduce susceptibility of HCC development (30). Furthermore jnk -/- mice and knockdown of JNK1 in human tumor cell lines displayed impaired tumor formation and cell proliferation, caused by reduced expression of MYC and increased expression of the CDK inhibitor p21 (26). In contrast, studies using MEFs from JNK1/2 knock-out mice have indicated that JNK is not only redundant for Ras-induced transformation and tumorigenesis in vivo, but may also have a tumor suppressive role by promoting apoptosis (27). Jnk1 -/- mice showed increased susceptibility to skin tumor formation whereas jnk2 -/- mice were more resistant to skin tumors, implying a tumor suppressive role of JNK1 but an oncogenic role of JNK2 in skin tumors (3132). JNK is also a key component of stress-induced apoptosis in MEFs (33). Furthermore, activation of the JNK pathway is critical in inducing cell death by DNA damage (3435) and has been associated with autophagy (3637).
The biological outcome of MAPK activation is dependent on the stimuli, the strength/duration of the signal and cell type/tissue specificity. For instance, transient JNK activation promotes cell survival, while prolonged JNK activation induces cellular apoptosis (38). This reflects a balance between the various upstream activators and negative regulatory mechanisms, which oppose pathway activation. The ability of the MAPK pathways to be activated by a variety of stimuli and to activate various downstream targets shows that they are important components in cell signalling and must be tightly regulated.
MAPK PHOSPHATASES (MKPs)
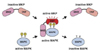
The magnitude and duration of MAPK activation determine the signaling output and are thus crucial in numerous biological processes (39). Abnormal MAPK signaling has been implicated in inappropriate cellular immune response and human malignancies. The MAPK phosphatase (MKP) or dual specificity phosphatase (DUSP) protein family, has been identified as major negative regulators of MAPKs (4041). MAPK inactivation occurs through dephosphorylation of threonine and/or tyrosine residues within the Thr-X-Tyr motif located in the MAPK activation loop (Fig. 2).
The MKPs share a common structure that comprises a C-terminal catalytic domain, which shares sequence similarity to the prototypic VH-1 DUSP encoded by vaccinia virus, and an N-terminal non-catalytic domain containing both a conserved cluster of basic amino acid residues involved in MAPK recognition (404243). MKPs are highly specific for MAPKs over other kinase substrates, but vary in their substrate specificities among the members of the MAPK family and subcellular localizations.
MKPs IN CANCER
The role of MKPs in cancer has been less well-defined than their associated MAPKs despite increasing evidence that disturbances in MKP function may play a role in cancer development. Studies have correlated loss of MKPs with cancer development or progression as a result of loss of negative regulation of MAPKs. However, gain of MKP expression has also been associated with cancer progression, drug resistance and poor patient prognosis (44).
Due to the emerging role of the MKPs in regulating cancer cell growth and death, there has been increasing interest in studies regarding the role of MKPs in cancer. This often begins with the establishment of a clinical correlation between tissue expression of a particular MKP, the activity of the corresponding MAPK substrate and effect on the disease. Following this, studies in cancer cells seek to correlate MKP expression and function to a tumor phenotype in vitro. Lastly, in vivo studies utilize mouse xenograft models to assess tumor formation and growth, or specific mouse models carrying one or more mutations or deletions of MKPs to examine specific types of cancers. However, despite this increase in MKP studies, the role of the MKPs in human cancers remains poorly defined.
ROLE OF MKP-1/DUSP1 IN CANCER
MKP-1 was the first MKP to be discovered and has been shown to be involved in various functions such as metabolic signaling (79), skeletal muscle function (80), inflammatory response (8182) and cancer (83). The study of MKP-1 has been the most advanced in the context of cancers compared to the other MKPs. Studies have revealed that MKP-1 was over-expressed in many cancer types such as breast (48), lung (45), prostate (84), ovarian (49), pancreatic (47), and gastric adenocarcinoma (Table I) (46). Clinical studies have also shown that MKP-1 expression is correlated with cancer progression and may be a useful prognostic marker. For instance, MKP-1 expression was found to be highly expressed in the malignant tissues of human breast cancer patients compared to non-malignant samples (8586). The increased expression of MKP-1 correlated with reduced JNK activity, suggesting that therapies that reduce the expression or activity of MKP-1 might enable the expression of the pro-apoptotic signaling by JNK in malignant cells. In pancreatic cancer, MKP-1 mRNA and protein levels are elevated, and down-regulation of MKP-1 expression in pancreatic cancer cells resulted in reduced anchorage-dependent and independent tumor growth (47). The increased levels of MKP-1 expression were found to have an inverse correlation with both JNK activity and apoptosis markers, indicating that MKP-1 might mediate anti-apoptotic effects in pancreatic cancer via its regulation of JNK activity. In HCC, decreased MKP-1 expression in tumor cells was found to be extensively correlated with improved disease-free survival rates, and reduced serum alpha-fetoprotein levels and tumor size (50). In primary ovarian tumors, moderate to strong expression of MKP-1 was detected in 57.6% of invasive ovarian carcinomas and MKP-1 expression was a prognostic marker for shorter progressionfree survival (49).
MKP-1 has also been assessed in relation to cancer therapy and resistance. In breast cancer, several other studies have indicated that MKP-1 may be an adverse prognostic factor in breast cancer patients and contributes to chemoresistance (858788). Studies have shown that MKP-1 overexpression lead to increased resistance to cisplatin in human lung cancer cell lines while knockdown of MKP-1 enhanced cisplatin-mediated apoptosis in the lung (89). Additionally, a recent study reported that MKP-1 translocates to the mitochondria after cell irradiation and inhibits the pro-apoptotic activities of JNK in breast cancer cell lines (86). In colorectal cancer (CRC), it has been shown that MKP-1 overexpression was associated with resistance to cetuximab-based chemotherapy in KRAS wild-type patients with metastatic CRC, indicating a role for MKP-1 as a negative predictive biomarker of response to cetuximab (90). In HCC cell lines, chemotherapy in combination with knockdown of MKP-1 resulted in significantly greater efficacy than chemotherapy alone (91). In lung cancer cell lines, silencing (siRNA) of MKP-1 expression resulted in enhanced activation of JNK and p38 by cisplatin compared to the parental cells, and this correlated with an increase in sensitivity to cisplatin (45). Cisplatin has been shown to induce MKP-1 in lung and ovarian cancer cell lines and this occurs via an ERK-dependent pathway (9293). The studies demonstrated that ERK is able to phosphorylate MKP-1, resulting in stabilization of the protein and resistance to ubiquitin-mediated degradation. MKP-1 over-expression also protected breast cancer cells from apoptosis triggered by doxorubicin, mechlorethamine and paclitaxel (94). Importantly, these studies show that MKP-1 conferred resistance to these chemotherapeutic agents by inhibiting JNK pathway (798994). Furthermore, knock-down of MKP-1 in breast cancer cell lines was able to promote tamoxifen-induced cell death via JNK (95). Interestingly, another DNA damaging agent, campthothecin, was found to induce apoptosis in the HCT116 CRC cell line by down-regulating MKP-1 expression and a reciprocal prolonged induction of ERK activation in the nucleus (96). This suggests that campthothecin treatment promotes antitumor effects by inhibition of MKP-1 activity on the ERK pathway. Together, these studies provide evidence of MKP-1 in mediating resistance to chemotherapy by regulating the MAPK pathways. However, the specific MAPK pathway regulated by MKP-1 in conferring chemo-resistance seems to be dependent on cell/tissue type and chemotherapeutic agent used. Overall, these findings suggest that MKP-1 may play a critical role in cancer development, may be a useful marker for predicting the survival of cancer patients and may be significant in determining treatment outcomes.
ROLE OF MKP-2/DUSP4 IN CANCER
Like MKP-1/DUSP1, MKP-2/DUSP4 is also predominantly localized in the nucleus (97). However, its involvement in cancer is less well-known compared to MKP-1. Genetic studies have provided evidence of increased mRNA levels of MKP-2 in hepatomas (53), pancreatic cancers (54), familial modulatory thyroid carcinoma (FMTC), multiple endocrine neoplasia (MEN), and papillary thyroid carcinoma (98). Moreover, MKP-2 expression was increased 3-fold in human breast cancer samples as compared to non-malignant samples where it appears to be co-expressed with MKP-1 (48). In pancreatic cancer cell lines harboring activating mutations in K-Ras, MEK-dependent over-expression of MKP-2 has been reported (54). There has also been evidence of MKP-2 over-expression in benign serous borderline tumors (SBT) in the ovary (44). The expression levels of MKP-2 were found to be higher in SBT compared with serous carcinomas (SCAs) and it has been implied that MKP-2-mediated suppression of ERK-dependent events associated with degradation of the extracellular matrix may be responsible for the more benign behavior of SBT (55). A recent study by Gröschl et al., discovered that MKP-2 was highly expressed in CRC with high frequent microsatellite instability (MSI-H) compared to microsatellite stable (MSS) CRC (99). Additionally, overexpression of MKP-2 in CRC cell lines led to increased proliferation, possibly by stimulating growth promoting genes such as Myc and cyclin D1 (99). MKP-2 was also found to be highly expressed in intestinal epithelial cells expressing oncogenic forms of KRAS and BRAF, and in KRAS and BRAF mutant CRC cells, with corresponding nuclear ERK inhibition (100). It is possible that sustained nuclear ERK activation is unfavourable for the cancer cells and thus the nuclear ERK phosphatases may be an alternative target in treating CRC displaying aberrant KRAS/BRAF/ERK signalling. In line with this, inhibition of MKP-2 attenuated the in vitro and in vivo proliferation of mammary tumor cells derived from a RET-MEN2A transgenic mouse by suppressing cyclin B1 expression, and leading to dysregulation of the cell cycle (98).
In contrast, several other studies have implicated MKP-2 as a negative regulator of cancer. Screening studies have suggested that MKP-2 may be a tumor suppressor/cancer susceptibility gene linked to chromosome 8p, which is frequently mutated in breast and prostate cancers (57). Low levels of MKP-2 expression also correlated with higher post neo-adjuvant chemotherapy cancer cell proliferation and a reduction in recurrence-free survival in breast cancer patients (101). Overexpression of MKP-2 in breast cancer cell lines reduced ERK activation and increased cell killing by docetaxel (101). In addition, EGFR-mutations have associated with loss of MKP-2 within the 8p locus in lung cancer (56). This is supported by another study that observed high rates of loss of heterozygosity (LOH) at markers adjacent to or within MKP-2 in breast cancer cell lines, supporting their status as candidate tumor suppressor genes (102). Furthermore, MKP-2 was found to be a common epigenetically silenced gene in glioma and overexpression of MKP-2 inhibited glioblastoma cell growth (58). More recently, MKP-2 was found to be induced by tamoxifen in breast cancer (MCF-7) cells and MKP-2 overexpressing MCF-7 cells have increased sensitivity to tamoxifen treatment by inhibiting ERK-mediated proliferation and tamoxifen resistance (103). Interestingly, expression of MKP-2 was reported to rescue 293T cells from apoptosis upon UV or cisplatin exposure by negative regulation of JNK activation but not through regulation of ERK (104). These findings indicate that MKP-2 may have both positive and negative regulatory roles in cancer development although the mechanisms are still poorly understood.
ROLE OF MKP-3/DUSP6 IN CANCER
MKP-3/DUSP6 is the best characterized of the cytoplasmic phosphatases and specifically targets ERK1/2 over the stress-related MAPKs (105). A tumor suppressive function of MKP-3 which is mainly through negative regulation of ERK has been supported by studies on various cancers including pancreatic cancer, lung cancer, nasopharyngeal carcinoma (NPC), breast cancer as well as head and neck cancer. For instance, Mkp-3 is localized on chromosome 12q21, one of the regions of frequent allelic loss in pancreatic cancer cell lines, suggesting that this molecule may play a role in cancer pathogenesis (61). In a majority of these cell lines, reduced levels of MKP-3 was accompanied by activating mutations in KRAS (61). MKP-3 expression was slightly upregulated in primary pancreatic cancer tissues and dysplastic in situ carcinoma cells, but down-regulated in invasive carcinoma, particularly in the poorly differentiated type (60). The loss of MKP-3 and gain-of-function mutations of KRAS may synergistically lead to activation of the ERK1/2 pathway, thus contributing to the development of the invasive phenotype (60). MKP-3 was also observed to be induced in myeloma cells harbouring a constitutively active mutant NRAS gene, and in human melanoma cell lines with activating mutations in BRAF (106107108). These studies suggest that the expression of MKP-3 in response to activated RAS or RAF might serve as a negative feedback to the RAS/RAF/MEK/ERK1/2 pathway.
In primary lung cancers, decreased MKP-3 expression was associated with increase in growth activity and histological grade of tumors. MKP-3 expression levels were also found to be weaker in most lung cancer cell lines compared to immortalized airway epithelial cells, and forced expression of MKP-3 was able to suppress the growth of these cancer cell lines (62). In support of this, siRNA knockdown in high-MKP-3-expressing lung cancer cells showed enhanced ERK signaling and cellular proliferation, whereas MKP-3 overexpression in low MKP-3-expressing lung cancer cells resulted in attenuated ERK activation, growth arrest and apoptosis (109).
In eosophageal squamous cell carcinoma (ESCC) and NPC, MKP-3 was found to be down-regulated in 40% of ESCC biopsies and in 75% of NPC biopsies compared to their normal counterparts (67). The study also showed increased ERK activity in ESCC and NPC cell lines expressing low levels of MKP-3 compared to immortalized eosophageal and nasopharyngeal epithelial cell lines, indicating negative regulation of ERK by MKP-3. On the other hand, MKP-3 overexpression in ESCC and NPC cell lines resulted in reduced cancer cell proliferation and migration/ invasion in vitro, as well as reduced tumor growth in vivo (67). The tumor suppressive function of MKP-3 in ESCC was further supported by a recent study showing that MKP-3 expression was negatively correlated to pathological grade and that exogenous MKP-3 expression in ESCC cell lines markedly increased apoptosis in vitro (110).
Interestingly, MKP-3 has been shown to be a ΔNp63α-targeted gene and important for ΔNp63α-mediated inhibition of cancer invasiveness and metastasis via inhibition of ERK (111). Knockdown of ΔNp63α resulted in reduced MKP-3 expression, increased ERK1/2 activation and increased cell migration and invasion which can be rescued by forced expression of MKP-3. In addition, increased expression of Rab2A, a small GTPase critical for vesicle trafficking, has been shown to be correlated with poor clinical outcomes in human breast cancer patients (112). It was found that Rab2A binds to ERK to protect active ERK from dephosphorylation by MKP-3. This in turn promotes the nuclear translocation of β-catenin and tumorigenesis of BCSCs in human primary breast cancer (112).
Furthermore, studies have suggested that MKP-3/DUSP6 is involved in chemoresistance via regulation of ERK activation. In ovarian cancer, forced expression of MKP-3 or treatment with an ERK inhibitor enhanced cisplatin sensitivity, whereas MKP-3 knockdown resulted in reduced cisplatin-induced cell death (64). In melanoma cells, MKP-3 overexpression prevented ERK-mediated induction of nucleotide excision repair proteins, ERCC1 and XPF, by cisplatin, resulting in increased sensitivity to cisplatin (113). In contrast, MKP-3 overexpression promoted resistance to cisplatin-mediated cell death in in vitro and in vivo models of glioblastoma, indicating that MKP-3 may function as a tumor promoter in these cancers (114). Since ERK activation has been shown to be responsible for mediating apoptosis in glioblastomas (115), this correlates with the chemo-protective function of MKP-3 in this cancer. This provides evidence that the role of MKP-3 is dependent on the function of ERK in triggering apoptosis or promoting resistance in response to DNA damaging agents.
ROLE OF MKP5/DUSP10 IN CANCER
MKP5 was originally identified as a phosphatase capable of regulating JNK and p38 MAPKs, but not ERK1/2, when overexpressed in cell lines (116117). However, studies using MKP5 deficient mice from our group demonstrated that this protein is able to dephosphorylate all the three groups of MAPKs as well as IRF3 in a cell/tissue and context-dependent manner (118). By regulating JNK in immune cells such as T cells, MKP5 plays a significant role in both innate and adaptive immune responses (119). In neutrophils, MKP5 selectively inhibits p38 activation and suppresses the expression of inflammatory mediators such as superoxide (120). In adipose tissue, increased p38 activation was detected in aged MKP5 deficient mice, which is correlated with the development of visceral obesity and adipose tissue inflammation in the mice (121). In intestinal epithelial cells, MKP5 was found to suppress ERK activation and cell proliferation (122). Moreover, this protein was found to directly interact and dephosphorylate IRF3 in macrophages in response to RNA virus infection, thereby inhibiting type I interferon responses (118). These studies demonstrate the important role of this molecule in various physiological and pathophysiological conditions.
MKP5 is emerging as an important player in cancer as well. In the HCC cell line HepG2, environmental carcinogens upregulate microRNA(miR)-181 to enhance p38 activation by targeting MKP5 (123). Inhibition of miRNA-181 led to enhanced expression of MKP5 and a corresponding suppression of p38 activation, which in turn reduced cancer cell migration. In pancreatic cancer cell lines, miRNA-92a targets MKP5 to promote JNK activation, thereby enhancing cell proliferation (124). These studies suggest that MKP5 is possibly a tumor suppressor in HCC and pancreatic cancer by inhibiting p38 or JNK.
Nomura and colleagues reported that MKP5 was frequently upregulated in CRC but not in lung carcinoma, breast cancer or glioblastomas (73). Intriguingly, they found that MKP5 functions as a scaffold protein and cytoplasmic anchor for ERK, preventing nuclear translocation of phospho-ERK, thereby inhibiting ERK-dependent transcription and cancer progression. Our study on MKP5 in intestinal epithelial cells and tumorigenesis demonstrated that this protein suppresses intestinal epithelial cell (IEC) wound healing and inhibits tumorigenesis by suppressing ERK-mediated signaling (122). We showed that loss of MKP5 in mice resulted in increased cell proliferation and wound healing due to upregulation of ERK activation and KLF5 expression. While these factors are important in intestinal healing and barrier function maintenance after injury, unregulated cell proliferation may result in cancer development. Indeed, MKP5 knockout mice developed more colon tumors with increased severity associated with increased intestinal epithelial cell proliferation and migration in response to dextran sodium sulphate (DSS)/azoxymethane (AOM) treatment compared to WT mice. In addition, overexpression of MKP5 in human CRC cells resulted in reduced tumor formation in immune deficient mice. Furthermore, high MKP5 expression is associated with better survival in CRC patients (122). These findings established MKP5 as a tumor suppressor in CRC, and a potential target for the development of therapeutic methods to improve CRC patient outcomes.
ROLE OF MKP7/DUSP16 IN CANCER
MKP7 (DUSP16) was identified as a phosphatase specific for p38 and JNK in vitro (125) although some groups have reported MKP7 to be specific only for JNK (126). However, using MKP7 deficient cells, we demonstrated that this protein inactivates ERK and JNK in T cells in response to T cell receptor stimulation to specifically regulate Th17 response (127). In addition to its function in immunity, a regulatory role of MKP7 in cancer has been implicated. For instance, studies have suggested that MKP7 may be downregulated in cancers by genetic or other mechanisms. In this respect, it is worth noting that the MKP7 gene is located on chromosome 12p, an area of frequent chromosomal loss in human tumors such as leukemia (74), NSCL cancer (75) and prostate carcinoma (76). Furthermore, MKP7 overexpression, through inactivation of the JNK pathway, has been shown to reduce Rat-1 cell growth and suppress transformation capabilities of BCR-ABL oncogene transformed cells in vitro and in vivo (74). Downregulation of MKP7 by miR-24, whose expression is induced by compromised RUNX function, has also been linked to development of acute myeloid leukemia (77). The reduction of MKP7 expression led to increased JNK and p38 phosphorylation, which in turn stimulated myeloid cell growth and inhibited differentiation. There was also evidence showing that MKP7 may regulate sensitivity of cancer cells to anti-cancer therapies. For instance, methylation-dependent transcriptional silencing of MKP7 in Burkitt's lymphoma cells, but not other B cell lymphomas, abrogated the negative regulation of JNK activity to enhanced the sensitivity of cells to chemotherapeutic agents that activate JNK, such as doxorubicin, sorbitol and cisplatin (78). Moreover, a study demonstrated that a novel anticancer agent, NSC-741909, induced ROS-mediated inactivation of MKP7 in NSCL cancer cell lines to activate the JNK signaling pathway and apoptosis (128).
OTHER MKPs IN CANCER
Several other MKPs, including PAC-1/DUSP2, MKP4/DUSP9, and MKP-X/DUSP7, have also been implicated in the development of cancers. For example, in ovarian carcinoma, high levels of PAC1 mRNA predicted significantly worse overall survival as compared to low expression (51). Interestingly, in acute leukaemia, loss of DUSP2/PAC-1 expression has also been associated with constitutive ERK activation (52), suggesting that PAC-1 may function as a tumor suppressor. For MKP4/DUSP9, a tumor suppressive function has been implicated. It was reported that loss of MKP4 was associated with squamous cell carcinoma (SCC) but not in benign papillomagenic lineages (69). Overexpression of MKP4 resulted in tumor cells microtubule disruption and cell death. Finally, MKP-X/DUSP7 has been found to be constitutively highly expressed in bone marrow and peripheral leukocytes from acute myeloid leukaemia (AML) and acute lymphoblastic leukemia (ALL) patients, whereas very low levels of MKP-X was detected in the bone marrow of healthy patients (68), suggesting the possible involvement of this molecule in the pathogenesis of leukaemia.
TARGETING MKPs FOR CANCER TREATMENT
Aberrant expression of various MKPs/DUSPs has been observed in human cancers. However, whether overexpression or loss of expression of the MKPs contributes to malignant phenotype depends on the specific MKP and cancer/tissue type. The use of genetic knockout mouse models for individual MKPs allows for the study of whether gain/loss of expression is associated with the development and progression of cancers. These studies also help in investigating the roles of MKPs in mediating the sensitivity of tumors to chemical and physical agents used in both conventional and novel cancer therapies. For example, MKP-1, MKP-3 and MKP7 have been linked to resistance to chemotherapeutic treatment thus providing possible targets for the design of new therapies.
Therefore studies have been done to identify phosphatase inhibitors specific for individual MKPs which may have anti-tumor properties. Sanguinarine was identified as a potent and selective inhibitor of MKP-1 in HeLa and PANC-1 cancer cell lines (129). Along with its analogue, chelerythrine, sanguinarine was able to inhibit MKP-1 in vitro and enhanced phosphorylation of ERK and JNK (129). Thus sanguinarine and its analogs may serve as novel MKP-1 inhibitors in the treatment of human cancers. Another MKP-1 inhibitor, triptolide, has been shown to enhance cisplatin-mediated apoptosis in osteosarcoma (130) and ovarian cancer cells (93). A group of uracil quinolones have also been identified by Arnold et al., from a chemical library of MKP-1 inhibitors which may be useful in studying the role of MKP-1 in cancers (131). Two of the pyrrole carboxamides, PSI2106 and MDF2085 have also been identified as selective inhibitors of MKP-1 but their role in cancer therapy has not been examined (132). In addition, using a high-content, fluorescence-based chemical complementation assay for MKP activity, Vogt and Lazo identified NSC95397, a compound which inhibited both MKP-1 and MKP-3 in vitro, and restored paclitaxel sensitivity in breast cancer cells exposed to dexamethasone (133). Thus the use of specific inhibitors targeting the MKPs may provide novel and specific therapies against cancers.
CONCLUSION
The MAPK protein family plays a diverse and at times contradictory role in the development of human cancers. As negative regulators of the MAPKs, the MKPs/DUSPs thus play important roles in cellular responses that are integral to cancer development and progression. Studies have suggested that the MKPs are potential biomarkers in cancers and MKP expression is often correlated with patient prognosis and chemotherapeutic sensitivity. However, the correlation between an MKP and a tumor phenotype may vary depending on the type of cancer. Further studies have to be conducted to discern the expression levels of the MKPs and how they affect the downstream signaling pathways and responses in human cancers. The understanding of how abnormalities in MKP expression and the regulation of the MAPK pathways by these phosphatases may thus be critical to developing more effective therapies against cancer.

 XML Download
XML Download