

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Dendritic cells (DCs) mediate innate and adaptive immunity of the immune system. A major role of DCs is to present antigens to T cells. As professional antigen-presenting cells (APCs), DCs activate T cells through co-stimulatory molecules such as CD80 and CD86. These are the classical immunogenic characteristics of DCs. In contrast, tolerogenic DCs (tolDCs) contribute to tolerance rather than immunity. TolDCs can induce T cell anergy and generate regulatory T (Treg) cells. TolDCs-mediated immune suppressive responses are achieved by the interaction between co-inhibitory molecules like programmed death ligand 1 (PD-L1) and ligand 2 (PD-L2) on DCs and programmed death 1 (PD-1) on T cells. In addition to these surface molecules, anti-inflammatory cytokines such as interleukin-10 (IL-10) and transforming growth factor beta (TGF-β) from DCs can promote tolerance. How are tolDCs generated or induced and what are the factors involved in this process? It is necessary to first distinguish tolDCs from immature DCs (imDCs). The capacity of imDCs for inducing an immune response is inferior to that of mature DCs (mDCs) because imDCs express a lower frequency of MHC and co-stimulatory molecules on their surface than mature DCs. In addition, imDCs have no ability to secrete pro-inflammatory cytokines such as IL-6 and IL-12 (1). Because of their immature phenotypes, imDCs have limited ability to prime T cells and induce an immune response and thus have been considered tolDCs. However, the phenotype and definition of tolDCs remain controversial.
Here, we define the typical phenotype and the characteristic of tolDCs based on two perspectives: expression of regulatory molecules and secretion of cytokines. In this sequence, we will sum up the various factors that have been widely known to induce tolDCs in vitro. Finally, the importance of tolDCs in clinical trials and their therapeutic application will be discussed in this review.
PHENOTYPIC AND FUNCTIONAL CHARACTERISTICS OF tolDCs
Expression level of co-stimulatory and inhibitory molecules on the cell surface of tolDCs
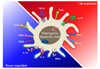
CD80 and CD86 on the surface of DCs can interact with CD28 on T cells. The interaction can trigger a signal to stimulate a T cell immune response. Therefore, DCs expressing a low level of co-stimulatory molecules may have a decided tolerance advantage.
On the contrary, surface inhibitory molecules such as PD-L1 and PD-L2 on DCs interact with PD-1 on T cells. The interaction between molecules results in anergy or functional inactivation of T cells (123). Although it has been reported that overexpression of PD-L1 or PD-L2 on DCs did not inhibit proliferation of T cells (4), high expression levels of PD-L1 or PD-L2 on the surface of DCs can still be a phenotypical characteristic of tolDCs, as shown in many individual studies (45678910111213). The expression of another death signal molecule CD95, a FAS-ligand, can also be a sign of tolerance (1415).
Cytokine-secreting pattern
In addition to the expression of surface molecules, a profile of cytokines secreted from DCs can represent the tolerogenic potential. The pro-inflammatory cytokine IL-12 produced by DCs stimulates the growth and function of T cells (16). Moreover, IL-12 can stimulate the secretion of interferon-gamma (IFN-γ) or tumor necrosis factor alpha (TNF-α) and can activate natural killer (NK) cells (16). Therefore, DCs that have a low ability to secrete such inflammatory cytokines can have 'tolerogenic potential'. DCs that secrete a high dose of anti-inflammatory cytokines such as IL-10 and TGF-β can also be considered tolDCs because IL-10 inhibits the synthesis and induction of pro-inflammatory cytokines such as IL-12, TNF-α, and IFN-γ (171819). Furthermore, IL-10 impairs the activation of NK and T cells (2021), and TGF-β is important for the maintenance and survival of Treg cells (22).
Other characteristics of tolDCs
It has been reported that the direct effect of IL-12 on DCs is to promote nuclear localization of nuclear factor kappa- light-chain-enhancer of activated B cells (NF-κB) (23). Therefore, a low expression level of NF-κB, which is widely related to the activation of DCs, could be another sign of tolerogenic characteristics of tolDCs.
Indoleamine 2,3-dioxygenase (IDO), an immune checkpoint molecule produced by tolDCs, is also related to the inhibition of T cell proliferation and survival (2425262728). Moreover, IDO has been reported to promote functional development of Treg cells (29). Thus, the induction or up-regulation of IDO in DCs can tolerate the immune environment of the host.
In summary, tolerogenic potential in DCs is indicated by the presence of down-regulation of factors related to boosting the immune response and up-regulation of inhibitory molecules, anti-inflammatory cytokines, and related enzymes (Fig. 1).
GENERATING tolDCs IN VITRO
Generation of DCs in vitro
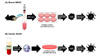
It is necessary to know the factors that are responsible for the induction of DCs with tolerogenic characteristics. Because of the scarcity of DCs and their various subsets in vivo, many in vitro DC studies have been performed. In a murine system, the classic method of investigating DCs is using bone marrow-derived DCs (BMDCs). BMDCs can be generated by 6~10 days culture in vitro (3031). In this system, DCs are differentiated from bone marrow stem cells, and the culture medium must contain granulocyte-macrophage colony-stimulating factor (GM-CSF) (32). After reaching one-third of the total culture period, fresh medium is added at the same volume as the pre-existing medium. After reaching two-thirds of the total culture period, half of the culture medium is replaced with fresh medium. At the end of the culture period, BMDCs can be harvested by collecting suspended cells. These harvested 'untouched BMDCs' are imDCs. Additional treatment of LPS, TNF-α, or IFN-γ causes imDCs to become fully mature DCs (Fig. 2A). Though the culture period and concentration of GM-CSF can vary, this method is highly standardized. The other technique for generating DCs, especially from humans, is differentiating DCs from peripheral monocytes (33343536). DCs differentiated in this system are called monocyte-derived DCs (MoDCs). Except that the origin of the DC is a monocyte, the other processes and principles of culture are almost identical to those of BMDCs (Fig. 2B). Incidentally, trials for differentiating human DCs from bone marrow have also been performed (3738).
Generation of tolDCs from DCs in vitro
When BMDCs or MoDCs are stimulated with certain factors, the DCs are able to turn into tolDCs (Fig. 3). While a variety of factors with the ability to induce tolDCs have been reported (39), there is no standardized protocol with the optimal concentration and treatment time. In the next section, we will introduce certain factors that are universally known as inducers of tolDCs.
VARIOUS FACTORS INDUCING tolDCs
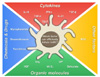
Cytokines
There are several cytokines known to induce tolDCs from BMDCs or MoDCs in vitro. Among them, the most well-known cytokines are IL-10, IFN-γ, and TGF-β. IL-10-treated BMDCs have a low capacity to secrete inflammatory cytokines and to prime T cells (40). Moreover, after IL-10 treatment, BMDCs are inhibited in their maturation (4142). The effects of IL-10 on BMDCs also correspond to DCs isolated from mouse spleen ex vivo (43). In addition to IL-10, IFN-γ has been demonstrated to induce tolerogenicity in DCs. IFN-γ-treated MoDCs cannot efficiently present alloantigens to T cells. In addition, when co-cultured with naïve CD4+ T cells, IFN-γ-treated MoDCs induce a high level of FoxP3 expression in CD4+ T cells and trigger the regulatory function of T cells (44). Notably, the destiny of DCs, whether they become suppressors or activators, depends on the concentration of IFN-γ (454647). High doses of IFN-γ inhibit maturation of MoDCs and induce their regulatory functions. For example, as the concentration of IFN-γ increases, the ratio of IL-10 to IL-12 produced by DCs increases but the cytotoxicity of CD8+ T cells that are co-cultured with the DCs decreases (4546). On the contrary, Kerkar and his colleagues reported that IFN-γ triggers differentiation of MoDCs and the expression of co-stimulatory molecules as well as the MHC I molecule (47). IFN-γ treatment, when added to monocytes in the early differentiation stage, often results in the induction of macrophages rather than DCs (48). However, after exposure to a low dosage of IFN-γ, MoDCs are differentiated to tolDCs, thereby hindering the T cell response (47). Taken together, the treatment time and the concentration of IFN-γ are crucial for generating tolDCs, which suggests that a more defined protocol is still needed. Another cytokine, TGF-β, when added at the final stages of BMDC differentiation, has been reported to down-regulate the expression of co-stimulatory molecules and inhibit the production of inflammatory cytokines in BMDCs (4049).
Organic molecules and chemicals
Dexamethasone (Dexa) is a steroid medication widely used for the treatment of various diseases. There are many reports showing that Dexa is a tolerogenic compound. Dexa-stimulated DCs tend to exhibit down-regulated expression of co-stimulatory and MHC molecules, and subsequently an impaired T cell immune response (405051525354). Furthermore, in vitro, Dexa treatment of DCs causes T cells to produce high amounts of IL-10 (54). While Dexa treatment methods for generating tolDCs differ among studies, Dexa is a tolerogenic reagent.
Vitamin D3 (VitD3) is another well-known tolerogenic reagent. The addition of 1,25(OH)2D3, an active form of VitD3, into DC culture results in the induction of tolerogenic surface molecules and the production of anti-inflammatory cytokines. In VitD3-treated DCs, the expression levels of CD80, CD86, and MHC II decrease and also the production of IL-6 and IL-12 are limited, compared to non-treated DCs (5556). Functionally, VitD3 contributes to the down-regulation of IFN-γ in DCs, thereby suppressing the T cell immune response (56). Because of their practical use and easy handling, Dexa and VitD3 have also been utilized in vivo (57) and are often used in combination (5859).
Others
Interestingly, DCs co-cultured with mesenchymal stem cells (MSCs) have immune-regulatory characteristics. In this system, DCs are overlaid on MSCs and cultured together (60). Under the influence of MSCs, DCs decrease IL-12 production and down-regulate expression of co-stimulatory and MHC molecules, resulting in the induction of tolDCs and T cell unresponsiveness (61). For this reason, there are quite a few trials using MSCs to suppress immune responses in vivo (6263).
THERAPEUTIC APPLICATIONS OF tolDCs
TolDCs are now being tested for their therapeutic use in various types of diseases where immune suppression is needed. Diversely generated tolDCs are used for the treatment of hyper-immune and autoimmune disorders such as rheumatoid arthritis (RA), multiple sclerosis (MS), and excessive tissue destruction and can be used to alleviate transplantation rejection.
TolDCs in clinical treatment for hyper- and autoimmune disorders
RA is the most representative and common autoimmune disease and has typical symptoms: synovitis and progressive cartilage destruction. In a murine system, the collagen-induced arthritis (CIA) model is widely used for the study of RA. Until now, common therapies for RA have included administration of immunosuppressive drugs and biomolecules even though these methods have fatal side effects causing general suppression of the host immune system (536465). Thus, adoptive transfer of tolDCs into a pathologic area can be a more effective treatment for RA patients. Adoption of tolDCs ameliorates inflammatory arthritis by inducing the production of anti-inflammatory cytokines and the generation of Treg cells, subsequently impairing the T cell immune response (666768697071727374757677). In addition to RA, other clinical trials targeting autoimmune disorders have recently adopted tolDCs. The disorders include MS (78), primary Sjögren's syndrome (59), immune thrombocytopenic purpura (79), diabetes (80), uveoretinitis (81), and experimental autoimmune encephalomyelitis (57).
Use of tolDCs in transplantation
TolDCs also play an important role in maintaining grafted organs (82838485). As in the treatment of autoimmune disorders, the use of immunosuppressive drugs has been a dominant treatment method for transplantation recipients. However, as mentioned above, the use of these suppressors often causes a breakdown of the host immune system. Recently, the administration of regulatory cells has been investigated (86). Although there is still a long way to go before achieving the ideal application, therapy using tolDCs is potentially the most promising and powerful tool to escape graft destruction in transplantation patients compared with therapy using other regulatory cells. As immunoregulatory APCs, tolDCs may be able to regulate and control other immune cells, which together give rise to tolerance. Although there have been many trials in animal transplantation models (blood, skin, heart, kidney, and pancreatic islet transplantations), clinical trials in humans have not been widely reported (87). It is obvious that injection of tolDCs can effectively suppress host immune system by inhibition of Ag-specific T cells and induction of Ag-specific Treg cells, but there are still many questions to be solved for safe and effective clinical goal:
Which tolDCs should be used: donor or recipient? Which time-point is appropriate for injecting tolDCs: prior-, peri-, or post-transplantation? Which route is more effective for injecting tolDCs: intravenous, intradermal, subcutaneous, or another route? What are the ideal dosage and frequency of administration (87)?
CONCLUSIONS AND PROSPECTS
We have discussed the generation, characteristics, and therapeutic applications of tolDCs. The definite phenotype and biomarkers for tolDCs have not been defined and opinions regarding tolDCs are diverse. Sometimes, even immature or semi-mature DCs are simply regarded as tolDCs. Furthermore, conditioning tolDCs is also very complicated because tolerance-inducing factors and manipulations for generating tolDCs are extremely varied.
TolDCs are a promising tool to suppress immune responses in autoimmune disorders and transplantations. However, additional research is needed for clinical trials. First, in the preparation stage of conditioned DCs, specific protocols to generate tolDCs suitable for each application must be established. Establishing the appropriate timing and intensity of growth factor and tolerogenic reagent treatment is crucial. Furthermore, the migration, and toxicity issue of tolDCs in the host after injection should also be considered. As another concern, the stability of administered tolDCs in vivo must be also considered for effective therapeutic applications. Various types of tolDCs generated with diverse factors mentioned above were known to keep stably their tolerogenic phenotype and function in vivo (538889), whereas there have been some reports showing that semi-mature DCs, which have been usually considered as tolDCs (909192), became immunogenic cells when they were administered to in vivo (9394). Even if there are a lot of things to be investigated, tolDCs could be powerful immune cells with immense potential for innovative clinical therapy in the future.
 XML Download
XML Download