

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Fatty acid-binding proteins (FABPs) are a family of cytosolic proteins with a molecular weight of 14 to 15 kDa that are involved in the intracellular lipid responses in cells (1). To date nine different FABPs have been identified and named according to the tissues in which they were first identified, for example, liver-type (L-), intestinal-type (I-), heart-type (H-), adipocyte-type (A-), epidermal-type (E-), and brain-type (B-) FABP (1). Among these, the first described FABP, liver-type FABP (L-FABP) is highly expressed in the liver (2~5% of cytosolic protein) as well as in the kidney, lung, pancreas and intestine (1), and a key regulator of hepatic lipid metabolism by influencing the uptake, transport, mitochondrial oxidation, and esterification of fatty acids (2).
Since FABPs lack a secretory signal sequence, these proteins are not detected in the blood stream under normal circumstances (34). Thus, some FABPs have been to be used as diagnostic markers of tissue injury: L-FABP for liver damage (567), I-FABP for intestinal injury (89), H-FABP for acute myocardial infarction and ongoing myocardial damage in heart failure (1011), and B-FABP for brain injury (12). Recent studies, however, have shown that some of FABPs can be secreted from cells without cell damage. L-FABP is detectable in bile in the absence of cellular injury (), H-, A-, and E-FABPs are present in milk (1415), and several recent reports have shown that A-FABP is secreted from adipocytes into the circulation or extracellular spaces (1617). Furthermore, it has become evident that the A-FABP after release from adipose tissue has crucial hormonal functions on liver, pancreas and heart (181920). As for the role of L-FABP, different studies have shown that urinary L-FABP in humans would be a useful clinical marker for acute kidney injury, and prediction and monitoring of the progression of chronic kidney disease (212223). Other reports suggested that serum L-FABP level may serve as an early biomarker for lung damage in a model of acute respiratory failure (24) and renal injury following kidney transplantation (25), and as a new diagnostic marker for detecting non-alcoholic fatty liver disease (26). In addition, several correlative studies suggest that L-FABP may also, like the A-FABP, function as an endocrine factor by showing that increased serum concentrations of L-FABP are associated with obesity, insulin resistance and high blood pressure (2728). However, little is known about the extracellular biological function of secreted L-FABP.
In the present study, we investigate whether L-FABP has a biological function in the blood and other human cells. In addition, we determined serum L-FABP levels in patients with end-stage renal disease (ESRD) on hemodialysis (HD), who have been known to have altered lipid metabolism including fatty acid.
MATERIALS AND METHODS
Materials
A549 lung carcinoma and THP-1 monocytic cell lines were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). A549 cells were maintained according to the instructions and THP-1 cells were cultured in RPMI 1640 medium containing 10% fetal bovine serum (FBS). Huh7 cells were maintained in DMEM with 10% FBS. Human IL-6 enzyme-linked immunosorbent assay (ELISA) kit was obtained from R&D Systems (Minneapolis, MI, USA).
Reverse transcription polymerase chain reaction (RT -PCR ) and molecular cloning for E. coli expression vectors
Total ribonucleic acid (RNA) was isolated with TRI Reagent® (Sigma-Aldrich, St. Louis, MO, USA) from the Huh7, HepG2, HT29, colo205, sw620, KM12C cells (not shown). A pair of human L-FABP sense primers, 5'-ctacagtggacagtctggtc-3', and reverse primer, 5'-gtttaaa ttctcttgctgattc-3', was used for the RT-PCR. Moloney murine leukemia virus-reverse transcriptase (Beams Bio, Korea) was used for converting 2 µg of total RNA to first strand complementary deoxyribonucleic acid (cDNA), and the PCR was performed at 94℃ for 45 s, 70℃ for 2 min, and 56℃ for 1 min for 30 cycles.
For E. coli expression vector, the PCR product of human L-FABP cDNA was ligated into T&A cloning vector (RBC, Taiwan). The cDNA of L-FABP was amplified by PCR with the sense primers of human FABP1 with 4 overhangs, NdeI site with artificial start codon for cloning into pET21a, 5'-ATATCATATGAGTTTCTCCGGCAA- 3', and the reverse primer of human FABP1 with 4 overhangs and XhoI site for cloning into pET21a, 5'-AT ATCTCGAGaattctcttgctgatTCT-3'. Inserts were digested with NdeI and XhoI and then transferred into pET-21a (Invitrogen, Carlsbad, CA, USA) for recombinant protein expression in E. coli.
Expression of recombinant protein
Human recombinant L-FABP protein was expressed in E. coli and Rosetta cells (Novagen, Madison, WI, USA) and purified by TALON affinity columns (Invitrogen) by using His6-tag at the N terminus of recombinant proteins. The TALON affinity-purified proteins were subjected to high performance liquid chromatography (HPLC, GE Healthcare) with a C4 column (Grace Vydac, Hesperia, CA, USA). The recombinant protein was subjected to 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) for silver staining. In addition, the eluents of various fractions and known amounts of bovine serum albumin (BSA) were subjected to 10% SDS-PAGE and visualized by silver staining. A standard curve was created with density measurement of BSA standards and used to calculate estimates of human recombinant L-FABP protein concentration.
Patients
ESRD patients had received HD in Jeju National University Hospital Dialysis Unit were studied age of 61.6±1.8 (mean±SEM) years. This study was approved by the institutional review board of Jeju National University Hospital and informed consent was obtained for the patients. The control group included age and gender matched healthy volunteers (n=63), renal diseases of the ESRD dialysis (n=75), and diabetes patients (n=87). The ESRD dialysis patients were dialyzed 2 or 3 times per week on Monday–Wednesday–Friday, Tuesday–ThursdaySaturday, or Tuesday–Saturday using high-flux membranes (dialysis filter surface area, 1.7~2.1 m2), and exhibited an equilibrated Kt/V of 2.07±0.23. The mean duration of HD was 63.4±7.1 months.
Blood samples collection and cell culture
Blood samples of healthy volunteers were taken after an overnight fast and collected in heparin- or ethylenediamine tetraacetic acid (EDTA)-coated tube. And then we compared the effect of the two commonly used anticoagulants on the ability of human whole blood cells to induce IL-6 upon human recombinant L-FABP or IL-1α stimulation. Blood samples of ESRD patients were drawn before the start of a routine HD treatment on Monday or Tuesday and used to measure the levels of serum L-FABP. A549 (2.5×105/well), THP-1 (1×106/well) and human whole blood cells (1:2 dilution with RPMI) were seeded in a 96-well plate with 200 µl volume and then were treated with various concentrations of human recombinant L-FABP for overnight.
Measurement of IL-6 and L-FABP
IL-6 levels were measured in cell culture supernatants by sandwich ELISA. For the detection of human L-FABP in serum, we developed monoclonal antibody against L-FABP from BALB/c mice immunized with the purified human recombinant L-FABP (not shown). The secondary antibody for L-FABP was conjugated to horseradish peroxidase with the use of succinimidyl 4-(N-maleimidomethyl) cyclohexane-1-carboxylate and a two-step sandwich ELISA method was carried out for determining L-FABP concentrations in serum. The detection limit was approximately 80 pg/ml.
RESULTS
Generation of human recombinant L-FA BP
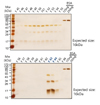
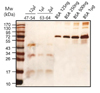
In order to purify human recombinant L-FABP protein, the E. coli extract was applied to a mini Talon affinity column. The primarily purified recombinant L-FABP protein was further purified by HPLC. The molecular size of His6-tag recombinant L-FABP was approximately 16 kDa. HPLC results indicated two groups of peak after 51 and 63 min (Fig. 1). Aliquots of the various fractions were resolved by 10% SDS-PAGE, and the protein bands were visualized by silver staining. A serious of protein bands with molecular weight of 16-, 32-, and 48 kDa were identified in nearly all fractions examined (Fig. 2A). Especially these protein bands were detected mainly in fractions 63 and 64 (Fig. 2B). We verified human recombinant L-FABP in the eluted fractions with a monoclonal antibody specific to human L-FABP. The anti-human L-FABP monoclonal antibody recognized the 16-, 32-, and 48-kDa human L-FABP (not shown). The results suggest that 16,32 and 48 kDa are the monomer, dimer and trimer forms of L-FABP, respectively (not shown).
In addition, the eluents from fraction 47 to 54 (12 µl) and from fraction 63 to 64 (1 and 2 µl, respectively), and known amounts of BSA were subjected to 10% SDS-PAGE and visualized by silver staining in order to estimate human recombinant L-FABP concentration (Fig. 3). A standard curve was created with density measurement of BSA standards and used to calculate estimates of human recombinant L-FABP protein concentration (not shown). Purified human recombinant L-FABP proteins from fraction 47 to 54 typically migrated as monomer, dimer and trimer, otherwise those from fraction 63 and 64 mainly migrated as monomer and dimer (Fig. 3).
Human recombinant L-FABP Stimulates production of IL-6 in A549 and THP-1 and human whole blood cells
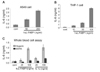
A549 and THP-1 cells were stimulated with various concentrations of human recombinant L-FABP for 24 h and then the induction of IL-6 was assessed by ELISA. The production of IL-6 was augmented along with the increased concentrations of human recombinant L-FABP in these cells, respectively (Fig. 4A and B). IL-6 production was also examined with human recombinant L-FABP or IL-1α treated human whole blood cells to investigate the biological activity of human recombinant L-FABP on human whole blood cells. As shown in Fig. 4C, human recombinant L-FABP or IL-1α stimulation increased IL-6 production. In addition, the effects of both human recombinant L-FABP and IL-1α were stronger in the human whole blood cells collected with heparin-coated tube than in those collected with EDTA-coated tube.
L-FABP concentration in human serum
We determined serum L-FABP levels using our ELISA assay in healthy control group and compared them with those in ESRD patients on HD because these patients have been known to have altered lipid metabolism including fatty acid (2930) and so might have altered serum levels of L-FABP. The serum levels of L-FABP were decreased in the ESRD patients on HD compared with those in the control group (2.7 ng/ml vs. 0.23 ng/ml, p<0.001) in Fig. 5. Also the level of diabetes patients L-FABP was lower than the level of normal control group. Especially, most of protein levels of ESRD patients on HD were lower than the lower limit of the control group (not shown).
DISCUSSION
FABPs were originally described as intracellular proteins that can affect lipid fluxes, metabolism and signaling within cells (1). However, numerous studies have shown the presence of FABPs in circulation (272831). In general, these circulating FABPs have been considered as markers of cell injury or death because they have no secretory signal peptide. However, a growing body of evidence suggests that the circulating form of FABPs have crucial hormonal functions in other tissues without cell damage. Several correlation studies have reported that elevated serum levels of A-FABP were associated with obesity, type 2 diabetes, hypertension, and cardiovascular diseases (2832), suggesting that there can be the link between circulating levels of A-FABP, and metabolic and cardiovascular disease. And a previous study reported that A-FABP directly depresses cardiomyocyte contractile activity after release from adipose tissue (18). Furthermore, another study revealed that exogenous A-FABP can induce replication and migration of human coronary artery smooth muscle cells (33). Moreover, some recent studies reported that exogenous A-FABP to primary hepatocytes can induce gluconeogenesis (19), and insulin resistance without altering beta-cell responsiveness in the pancreas (20). Altogether, previous data suggest that A-FABP has some hormonal functions on other tissues after release from adipose tissue. As for other FABPs, a few studies suggest that some FABPs such as L-FABP and E-FABP may have roles in systemic metabolism and cardiovascular system by showing the association between their serum concentrations, and metabolic and cardiovascular regulation markers (27283435). However, it has not been investigated, to our knowledge, that whether the secreted form of L-FABP is biologically active. In order to investigate its biologic activity, if any, we stimulated A549 lung carcinoma, THP-1 monocytic and human whole blood cells with human recombinant L-FABP and measured IL-6 with ELISA, and showed that IL-6 was induced in a dose dependent manner in these cells. Especially human recombinant L-FABP induced IL-6 in whole blood cells similar with IL-1α known as representative inflammatory cytokine. These results suggest that L-FABP in circulation or extracelluar space may have a role as a mediator of inflammatory process although we could not find out the mechanism by which L-FABP enters the blood compartment and if cell surface molecules that interact with L-FABPs on blood cells exist.
Additionally we found that the effects of both human recombinant L-FABP and IL-1α were potent inducer of IL-6 in the human whole blood cells collected with heparin-coated tube than in those collected with EDTA-coated tube. EDTA has an anticoagulant effect in blood by chelating calcium which is necessary for a wide range of enzyme reactions of the coagulation cascade and has been frequently used as the anticoagulant of choice for investigating hematological parameters, as cell morphology is preserved best over time (36). However, the use of heparin is recommended as the anticoagulant of choice in whole blood cell experiments because its influence on immune cell functions has been lesser than other anticoagulants, and anticoagulants acting by the chelation of calcium ion such as EDTA can remove the role of calcium in the activation of immune cells (373839). Thus, our results support that removal of calcium ion by EDTA can result in lower cytokine expression capacity in the whole blood compared with the use of heparin.
It has been shown that elevated serum levels of several FABPs are associated with lower glomerular filtration rate in general population and patients with angina pectoris, suggesting that FABPs are eliminated from the circulation mainly by renal clearance (2840). In addition, some studies reported that serum concentrations of H-FABP or A-FABP are increased in ESRD patients on HD compared to those in control subjects with normal renal function (41424344). However, there are only 2 published studies that investigated L-FABP levels in chronic kidney disease (CKD). One study reported that serum L-FABP levels in patients with CKD were higher than those in healthy volunteers (5.4±4.4 ng/mL vs. 1.2±0.7 ng/mL), and correlated with serum creatinine, suggesting that it may accumulate in the circulatory system through a decrease in glomerular filtration rates (21). However, the renal function of CKD patients in their study was relatively mild (serum creatinine, 1.1±0.8 mg/dl), so it may not represent the serum concentrations of L-FABP in ESRD patients. The other recent study showed that serum L-FABP levels of ESRD patients on HD (n=67) were higher compared with those of historical healthy control group (17.9±3.1 ng/ml vs. 1.2±0.7 ng/ml) and that serum L-FABP can serve as a biomarker for renal injury after kidney transplantation and to predict graft recovery and the need for HD (25). However, they did not compare the concentrations of L-FABP of ESRD patients with those of their own healthy control group. Contrary to the results of previous two studies, serum levels of L-FABP in ESRD patients on HD were significantly lower than those in the healthy control group in our study. So far there is no study to investigate the underlying mechanisms of increase or decrease in serum L-FABP levels in ESRD patients. Thus, the effect of ESRD on serum levels of L-FABP in patients remains unresolved. However, some studies reported that liver in rats with advanced CKD induced by 5/6 nephrectomy exhibits reduced nuclear peroxisome proliferator-activated receptor-α (PPARα) and downregulation of L-FABP which is upregulated by PPARα ligands (3045). Moreover fibrates stimulate cellular fatty acid uptake and catabolism by the beta-oxidation pathways through PPARα activation which are main roles of L-FABP in the liver (46). Based on these reports, we speculated that reduced activities of PPARα and L-FABP genes induced by ESRD may cause the reduced serum levels of L-FABP in ESRD patients.
In conclusion, we showed the biological activity of human L-FABP by demonstrating that its human recombinant form induces IL-6 production in human cell lines and whole blood cells. And the similar effect of L-FABP with IL-1α on whole blood cells indicates that circulating or extracellular form of L-FABP, if any, can be a mediator of systemic inflammation. In addition, we also found that the serum levels of L-FABP in ESRD patients on HD were significantly lower than those in the healthy control group, suggesting that severe renal dysfunction may affect the decreased induction of L-FABP in the liver.


 XML Download
XML Download