

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Adipose tissue is composed of adipocytes, stromal vascular fraction such as preadipocytes at various stage of differentiation, fibroblasts, vascular endothelial cells and heterogeneous population of immune cells (1). The obese state is now acknowledged as the main contributor to chronic low-grade inflammation (2). Recently, obesity has become a global epidemic and poses a large health burden as it is associated with excess adiposity that leads to an array of comorbidities such as type 2 diabetes, stroke, coronary heart disease and cancer (34). Obesity is an abnormal increase in adipose tissue mass leading to chronic inflammation and dysregulated adipocytokines such as proinflammatory mediators (CXCL8, IL-1, IL-6, TNF), angiogenic factors (VEGF) and metabolic regulators (leptin, adiponectin) (5). Interestingly, adipose tissue mass has an architectural organization in which metabolic cell such as adipocytes are in close proximity to macrophages, creating continuous interactions between immune and metabolic responses (5). It is now established that this interface contributes to the inflammatory response associated with obesity (6). Adipose tissue, a biologically-active endocrine tissue, serves as energy storage and secretes various proinflammatory cytokines and chemokines (7). Growing evidence suggests that cytokine and chemokine synthesis by human primary adipocytes may play a role in systemic inflammation, which describes the obese state (89).
Chemokines are chemotactic cytokines that instigate chemo-attraction, thus proinflammatory chemokines are produced in response to inflammation (10). There are four classified groups of chemokines according to the location of the conserved cysteine residues: XCL, CCL, CXCL and CX3CL, where X is any amino acid residue (11). Chemokines bind specifically to chemokine receptors which are seven-transmembrane spanning G proteincoupled cell-surface receptors followed by their own functional roles via the intracellular signaling pathway such as immune surveillance and inflammation (12). In this study, we used human preadipocytes and adipocytes and identified the profiles of chemokine signature, providing a better understanding into the complex interactions of the chemokine network between immune cells and adipocytes implicated in the pathogenesis of obesity-derived chronic low-grade inflammation.
MATERIALS AND METHODS
Reagents
The PCR array for customized human chemokines (CAPH10008) and SYBR® Green Master Mix (cat #: 330503) were purchased from SABiosciences/Qiagen (Frederick, MD, USA). Specific PCR primers for chemokines were obtained from Eurofins MWG Operon (Huntsville, AL, USA). All liquid culture media such as FBS (cat #: 26140), calf serum (cat #: 16010159), antibiotic-antimycotic solution (cat #: 15240096), and DMEM (cat #: 11965) were acquired from Invitrogen/Thermo Fisher Scientific Inc. (Waltham, MA, USA).
Human preadipocytes and adipocytes
The human preadipocytes and adipocytes were purchased from Cell Applications Inc. (San Diego, CA, USA). Preadipocytes and adipocytes were cultured in preadipocyte basal medium and adipocytes maintenance medium, respectively, according to manufacturer's instructions. Human preadipocyte and fully differentiated adipocytes were visualized under a digital inverted microscope (EVOS, Advanced Microscopy Group, Bothell, WA, USA) at 20 X magnification.
PCR array and qRT-PCR
After isolating total RNA and eliminating genomic DNA, the expression of chemokine ligands and receptors were analyzed using customized human chemokine PCR array (SABiosciences/Qiagen) on a Bio-Rad CFX96 real-time PCR instrument (Hercules, CA, USA). Briefly, reverse transcription reactions were performed at 42℃ for 15 min followed by 94℃ for 5 min. According to manufacturers' instructions, a real-time PCR was performed using a Bio-Rad CFX96 under the following two-step cycling program: 1 cycle at 95℃ for 10 min, 40 cycles at 95℃ for 15 sec and at 60℃ for 1 min. Data analysis was performed using PCR Array Data Analysis Software (http://www.sabiosciences.com/RTPCR.php) provided by SABiosciences/Qiagen. Changes in chemokine expression were confirmed using specific PCR primer sets described in Table I Values for PCR array are the means from duplicate experiments, while experiments for qRT-PCR were performed at least in triplicate.
Enzyme-linked Immunosorbent Assay (ELISA)
The level of secreted CXCL14 from conditioned media of preadipocytes and adipocytes media was assessed by enzyme-linked immunosorbent assay (ELISA) kit from Ray Biotech (Norcross, GA, USA) according to the manufacturer's instructions. The optical density of each well was determined using a microplate reader with 450 nm of wavelength. Assays were calibrated using serial dilutions of recombinant human CXCL14 (Ray Biotech).
Data analysis from GEO dataset
Data analysis was performed using microarray data sets deposited in the NCBI Gene Expression Omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo/) database under accession number GSE44000. Raw microarray data for chemokine network were RNA expression levels in subcutaneous adipose biopsies obtained from 7 lean (BMI<25) and 7 obese (BMI>30) postmenopausal women closely matched for age and NCEP III metabolic syndrome parameters exclusive of waist circumference (13). We employed Gitools 2.2.3 (http://www.gitools.org) based on Oracle Java 7, an open-source tool to perform Genomic Data Analysis and Visualization as interactive heat-maps (14).
RESULTS AND DISCUSSION
Obesity is known to alter metabolic and endocrine functions in adipose tissue followed by obesity-associated comorbidities. Inflammation in adipose tissue appears to be a key mechanism to explain the development of insulin resistance, as an important factors that leads to adipose tissue dysfunction (2). Cumulative evidence suggests that chemokines are one of important factors implicated in development of pathophysiological processes linking obesity and insulin resistance (10). In the present study, we characterized profiles of the chemokine network in human preadipocytes and adipocytes.
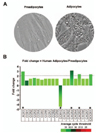
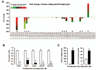
Morphologically, human preadipocytes revealed long, thin and flatted cells without detectable fat organelles while the matured human adipocytes had lipid accumulation in spherical form (Fig. 1A). Overall expression levels of chemokine receptors were absent or low (Fig. 1B), wherein adipocytes decreased CCR10 and increased XCR1, CXCR3, CXCR5 and CX3CR1 compared to preadipocytes (Fig. 1B). Our chemokine receptor signature indicates that chemokine receptors are unlikely to contribute to adipogenesis from preadipocytes to adipocytes. Meanwhile, there were significant differences in the chemokine signatures between preadipocyte and adipocyte. The dominant chemokines of preadipocytes were CCL2, CCL7-8, CCL11, CXCL1-3, CXCL6 and CXCL10-11 (Fig. 2A and 2B), which appears to be downregulated during adipogenesis. Interestingly, adipocytes expressed dominantly CXCL14 (Fig. 2A and 2C), indicating its important role in adipogenesis. In parallel, the ELISA data also revealed significantly higher protein level of CXCL14 in conditioned media of adipocyte compared to preadipocyte (Fig. 2C). Based on GEO dataset, chemokine signature in adipocytes obtained from healthy lean and obese postmenopausal women revealed that adipocytes from obese women had significantly higher levels in the following chemokines: CCL2, CCL13, CCL18-19, CCL23, CCL26, CXCL1, CXCL3 and CXCL14, compared to those from lean women (Fig. 3A and 3B). In particular, CXCL14 appeared to be expressed the highest levels in adipocytes from both lean and obese women (Fig. 3B). Based on the significant increase of CXCL14 in obese women compared to lean women (Fig. 3B), further study requires clarifying the positive relationship between CXCL14 levels and the degree of obesity to confirm CXCL14 as a biomarker for obesity.
Studies on adipose tissue in obese mice demonstrated increased level of CXCL14 (1516). High fat diet-fed CXCL14-deficient mice showed impaired white adipose tissue macrophage immobilization (17), suggesting the involvement of CXCL14 in macrophage recruitment in obesity. CXCL14 might recruit macrophages into adipose tissue, thus producing a wide range of cytokines, chemokines and growth factors to induce chronic inflammation that promotes obesity-associated diseases such as diabetes and cancer progression. Identification and characterization of still unknown CXCL14 receptor will be helpful in understanding the functional role and regulation of this adipocyte-secreted CXCL14 in obesityassociated comorbidities. In spite of downregulation of CCL2, CXCL1 and CXCL3 during adipogenesis (Fig. 2A), obese adipose tissues upregulated these chemokines (Fig. 3B). Because these chemokines are mainly the NF-κB driven chemokines (181920), obesity may contribute to a proinflammatory microenvironment by potentiating the NF-κB driven chemokines via recruitment of immune cells. This study identifies the chemokine signature during adipogenesis that could lay groundwork for the development of efficient therapeutic strategies based on the prevention of low-grade inflammation, which defines the obese state. Our finding suggests a new plausible marker to restore chronic, low-grade inflammation, by suggesting a distinct functional role of CXCL14 from other proinflammatory chemokines.


 XML Download
XML Download