

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cancer Immunotherapy targeting enhancement of anti-tumor T cell reactivity has received significant attention in recent years, mainly due to the favorable clinical responses of immune checkpoint inhibitors, such as anti-CTLA4 and anti-PD-1 antibodies (1). Anti-PD-1 antibodies in particular showed significant objective responses in clinical trials for some solid tumors, such as malignant melanoma (30%), non-small cell lung cancer (NSCLC; 20%), renal cell carcinoma (17%), and etc (2345). To this end, two PD-1 antibody drugs, nivolumab and pembrolizumab, have received US FDA approval for treatment of advanced melanoma, NSCLC, and metastatic renal cell carcinoma. Also, antibodies against PD-L1, a ligand for PD-1, are being tested in clinical trials for various types of cancers (6). Thus, therapeutic strategies targeting the PD-1/PD-L1 axis have considerable potential to contribute to the treatment of multiple types of tumors.
PD-1 is an inhibitory receptor expressed mainly on activated T cells, B cells and monocytes (78). PD-L1 and PD-L2 are best-known ligands for PD-1 (910). Both are expressed on hematopoietic cells (dendritic cells, mast cells, macrophages, T cells and B cells), whereas PD-L1 is also expressed on non-hematopoietic cells (endothelial and epithelial cells) (1112). Interestingly, PD-L1 expression is increased on various types of cancer cells, suggesting that this molecule is used for immunoresistance of cancer cells (13). Biochemically, PD-1 delivers inhibitory signal through its intracellular cytoplasmic domain, which contains immunoreceptor tyrosine-based switch motif (ITSM) and immunoreceptor tyrosine-based inhibitory motif (ITIM). Ligation of PD-L1 with PD-1 induces phosphorylation of ITSM and ITIM and recruits SHP-1 and -2, which inhibits T cell receptor downstream signaling (1415).
In murine in vivo studies, PD-1 deficiency led to the development of autoimmune diseases such as lupus-like syndrome and dilated cardiomyopathy (1617). Therefore, one can expect that systemic treatment with PD-1 blocking antibodies in cancer patients will lead to autoimmune side effects. Indeed, 10 to 15% of treated patients developed grade 3~4 drug-related toxicities, although these toxicities were less severe than those of blocking antibodies against CTLA4, another co-inhibitory receptor on T cells (181920).
In this study, to utilize PD-1 blockade in a T-cell specific manner rather than systemically, we tried to inhibit endogenous PD-1 function in T cells by overexpressing a PD-1 mutant on T cells that is designed compete with endogenous PD-1 in a dominant negative manner. The mutant receptor was generated by deleting the cytoplasmic domain of PD-1, which we call PD-1 decoy. T cells expressing PD-1 decoy showed increased production of IFN-γ when co-cultured with PD-L1 expressing tumor cells in vitro and showed increased tumor regression in vivo. Thus, inhibiting PD-1 function in tumor-specific T cells will be a beneficial strategy for enhancing T cell-mediated anti-cancer therapy without the systemic toxicity induced by PD-1 blockade.
MATERIALS AND METHODS
Mice and cell preparation
B6, OT-I and Pmel-1 mice were obtained from the Jackson laboratories. The mice were maintained in a specific pathogen-free facility at the Research Institute National Cancer Center, Korea in accordance with the guidelines of the Institutional Animal Care and Use Committee. E.G7 lymphoma cells and B16-F10 (B16) melanoma cells were purchased from ATCC. Phoenix Eco and Phoenix GP cell lines were provided by Garry Nolan (Stanford University).
Plasmid constructs
The cDNA of extracellular and trans-membrane portion of mouse PD-1 (amino acid 1-199) was cloned into pMIG-w retroviral expression vector (a gift from Yosef Refaeli, National Jewish Medical and Research Center) to generate PD-1 decoy. For PD-1-CD28 chimera, the cDNA of the cytoplasmic portion of mouse CD28 (amino acid 177~218) was linked to the cDNA of extracellular and trans-membrane portion of PD-1 (amino acid 1~199) by blunt end ligation. PD-1 decoy and PD-1-CD28 chimera cDNAs were inserted in front of the IRES-GFP cassette in pMIG-w to use GFP as an expression marker. All sequences were confirmed by automated DNA sequencing.
Production and transduction of retrovirus
Phoenix GP cells were transiently transfected with a VSV-G (pMD.G)-coding plasmid and the retroviral plasmids by calcium- phosphate transfection. After 48 hours, the cultured supernatant containing VSV-G pseudotyped retrovirus was harvested. Subsequently, Phoenix Eco cells were transduced with the supernatant overnight. After 3~5 days, GFPhi populations were sorted by flow cytometry to generate stable cell lines producing the retrovirus. The supernatants harvested from the stable cell lines were concentrated 10 fold using a 100 kDa cut-off centricon (Millipore) to increase T cell-transduction efficiency. B6 splenocytes were activated with anti-CD3 antibody (5 µg/ml) plus anti-CD28 antibody (2 µg/ml) for 24 hours. Splenocytes from OT-I and Pmel-1 mice were activated with OVA257-264 peptide (1 µM) or hgp100 peptide (1 µM), respectively. The activated T cells were transduced with the concentrated retroviral supernatant plus polybrene (6 mg/ml) by spin infection. After 48 hours, the transduced T cells were rested for 3 days in the presence of recombinant human IL-2 (30 U/ml) without further stimulation. More than 95% of live cells were T cells after the resting. Transduction efficiency was measured by GFP positivity using flow cytometry. PE-conjugated anti-mouse PD-1 antibody (Clone: J43, BD Bioscience) was used to detect PD-1 decoy, PD-1-CD28 chimera, and endogenous PD-1.
In vitro cell culture experiments
GFP positive retrovirus-transduced B6 splenocytes were sorted by flow cytometry. GFPhi populations were composed of 60~70% of CD8 T cells and 25~35% of CD4 T cells before cell sorting. The sorted T cells (2×104) were stimulated with indicated amount of anti-CD3 antibody in the presence of irradiated splenocytes (2×105) for 48 hours followed by IFN-γ ELISA. Retrovirus-tranduced OT-I cells (serial dilution from 105 to 102 cells) were cultured with MC38-OVA cells (1×104) or E.G7-OVA cells (1×105) for 24 hours. The cultured supernatants were harvested and IFN-γ was measured by ELISA. For testing the efficacy of PD-1-CD28 chimera, Pmel-1 cells were transduced and co-cultured with IFN-γ (20 ng/ml)-treated B16 melanoma cells for 48 hours followed by IFN-γ ELISA.
In vivo tumor regression model
E.G7 cells (2×106) were subcutaneously injected into B6 mice on day 0. After 7 days, the retrovirus-transduced OT-I cells (2×106) were adoptively transferred into the tumor-bearing mice via intravenous injection. Tumor growth was measured every 3 to 4 days from day 7 until mice were euthanized. The approximate tumor sizes were calculated using the following formula: length×width×π (mm2). When tumor sizes exceed 500 mm2, the mice were euthanized. Statistical comparisons were made using the Wilcoxon matched pairs test.
RESULTS AND DISCUSSION
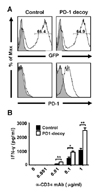
In order to generate a dominant negative mutant of PD-1, we designed a deletion mutant of PD-1, PD-1 decoy, which includes the extracellular and transmembrane domain of PD-1 with its intracellular domain deleted. This design allows PD-1 decoy to bind the ligand, but prevents it from delivering inhibitory signals inside the cell. Therefore, this mutant receptor is expected to compete with endogenous PD-1 for ligand binding and inhibit endogenous PD-1 function. To overexpress PD-1 decoy on T cells, we constructed a retroviral expression vector of this receptor. Retrovirus-transduced T cells were identified by GFP expression since the retroviral vector contains GFP cDNA as a reporter. When activated mouse splenic T cells were transduced with the retrovirus, transduction efficiency was approximately 65%, as measured by GFP positivity. GFP-positive T cells transduced with a PD-1 decoy-encoding virus were more strongly stained with anti-PD-1 antibody than those transduced with empty virus, which ensured overexpression of transduced PD-1 decoy (Fig. 1A). We hypothesized that overexpressed PD-1 decoy would diminish the co-inhibitory function of endogenous PD-1 and enhance functional activation of T cells. To test this idea, we sorted GFP-positive T cells via flow cytometry and stimulated them with anti-CD3 in the presence of irradiated splenocytes. When we measured secreted IFN-γ in the culture supernatant, PD-1 decoy-expressing T cells produced 2~3 fold more cytokines than control T cells (Fig. 1B). Therefore, it is very likely that PD-1 decoy interrupts binding of endogenous PD-1 to PD-1 ligands on splenocytes and inhibits endogenous PD-1 function.
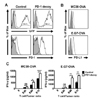
Next, we examined whether this functional enhancement of T cells could be applied to the anti-tumor reactivity of tumor-specific T cells. For this purpose, we utilized ovalbumin (OVA) as a model tumor antigen. OVA-transfected tumor cell lines and OVA-specific TCR-transgenic CD8 T cells (OT-1) served as target tumors and tumor-specific T cells respectively. OT-I T cells transduced with the PD-1 decoy-encoding retrovirus showed 55~65% transduction efficiency (GFP positivity), and the level of PD-1 decoy expressed in the GFP-positive population was higher than that of endogenous PD-1 expressed in the empty-vector-transduced control cells (Fig. 2A). For OVA-expressing tumor cell lines, we chose MC38-OVA, mouse colon cancer cells transfected with OVA, and E.G7, a mouse lymphoma cell line transfected with OVA because both cell lines express PD-L1, a ligand of PD-1, on their cell surface (Fig. 2B). When PD-1 decoy-transduced OT-1 T cells were incubated with these cell lines, they produced a higher amount of IFN-γ than control OT-1 T cells (Fig. 2C). The degree of enhancement of IFN-γ production was higher for E.G7-stimulated T cells than that for MC-38-OVA-stimulated T cells, which was correlated with higher PD-L1 expression on E.G7 cells than that on MC-38-OVA. Hence, PD-1 decoy improves the reactivity of tumor antigen- specific T cells in response to PD-L1-expressing tumor cells.
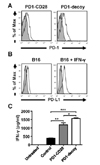
In our previous study on CTLA4, another inhibitory receptor on T cell surface, we reported that replacing the inhibitory cytoplasmic domain of CTLA4 with the stimulatory cytoplasmic domain of CD28 resulted in conversion of the negative signal of CTLA4 to a positive signal (21). Thus, overexpression of this chimeric receptor on tumor-specific T cells led to enhanced T-cell mediated tumor regression owing to the dominant negative effect on endogenous CTLA4 as well as the additional surrogate CD28 costimulatory signal. In this study, we also tested if a similar substitution of the CD28 cytoplasmic domain for the cytoplasmic domain of PD-1 would further increase T cell reactivity by providing additional costimulation. For this experiment, we generated a PD-1-CD28 chimera that consists of the extracellular and transmembrane domain of PD-1 and the intracellular cytoplasmic domain of CD28 (Fig. 3A). In this case, we used another tumor antigen-specific TCR transgenic (Pmel-1) CD8 T cells which recognize gp100 melanoma antigen. As a target tumor cell line, B16 melanoma cells were used. Although B16 cells showed moderate levels of PD-L1 expression on the cell surface, IFN-γ treatment further enhanced PD-L1 expression (Fig. 3B). Therefore, we incubated Pmel-1 T cells transduced with PD-1 decoy or PD-1-CD28 chimera virus with IFN-γ-treated B16 cells, and measured IFN-γ production from this co-culture. Similar to the results from OT-1 T cells, PD-1 decoy-transduced Pmel-1 T cells produced a higher amount of IFN-γ than control cells. PD-1-CD28 chimera also showed an enhancing effect on IFN-γ production from Pmel-1 T cells. However, it did not further enhance IFN-γ production from Pmel-1 T cells when compared with PD-1 decoy (Fig. 3C). This observation indicates that this chimera only exerted its dominant negative function without further costimulatory activity. This result is in stark contrast to a previous report that a PD-1-CD28 chimera in humans delivered CD28 signal in a CD28-null lymphoma cell line (22). This discrepancy may be due to slight differences in construct design between the two chimeras. Our chimera used the transmembrane domain of PD-1 molecule and the other used the transmembrane domain of CD28. Structurally, PD-1 exists as a monomer, whereas CD28 functions as a dimer. Thus, the CD28 domain in our construct may not have been able to dimerize to deliver its activation signal. In contrast, the previously published construct can dimerize because it has the transmembrane and cytoplasmic domain of CD28, which can be linked by disulfide bonds. This dimerization did not seem to affect the binding of this molecule to PD-L1. Nonetheless, our result showed that the inhibition of endogenous PD-1 signals with PD-1 decoy alone was sufficient to enhance the functional activity of tumor-specific T cells.
Finally, we investigated whether PD-1 decoy-mediated functional enhancement of T cells could increase therapeutic efficacy in vivo using a mouse model of anti-tumor T cell therapy. OT-1 T cell-mediated regression of OVAexpressing tumors is such a model. When OT-1 T cells were adoptively transferred to subcutaneous E.G7 tumor-bearing mice, tumor growth was significantly reduced. PD-1 decoy overexpression in OT-1 T cells greatly potentiated tumor regression in this model, consistently with the results from in vitro functional studies (Fig. 4). This result demonstrates that functional blockade of PD-1 in adoptively transferred tumor-specific T cells without systemic blockade of PD-1 may be a good strategy to enhance the efficacy of T cell therapy and to bypass potential side effect of autoimmunity at the same time.

 XML Download
XML Download