

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
cDC
conventional dendritic cell
CHR
contact hypersensitivity response
DC
dendritic cell
DTR
diphtheria toxin receptor
HDM
house dust mite
HMGN1
high-mobility group nucleosome-binding protein 1
ILC
innate lymphoid cell
LN
lymph node
MC
mast cell
moDC
monocyte-derived dendritic cell
PAMP
pathogen-associated molecular pattern
PDL2
programmed death ligand 2
TSLP
thymic stromal lymphopoietin
INTRODUCTION
Dendritic cells (DCs) bridge innate and adaptive immunity by activating antigen-specific CD4+ T cells via the provision of three signals. Signal 1 comprises the antigen-specific stimulation of T-cell receptor (TcR) through MHC class II together with an antigen-specific peptide complex. Signal 2 comprises costimulation mediated by the interaction of costimulatory molecules on DCs (e.g., CD80 and CD86) and their ligands on T cells (e.g., CD28), and is crucial for the clonal expansion of TcR-stimulated T cells. Signal 3 is mediated by cytokines provided mainly by activated DCs (1) and triggers the polarization of naïve T cells into effector T cells.
All of these signals 1, 2, and 3 contribute to the polarization of naïve T cells to effector T cells; however, the type of STAT activation in T cells triggered by signal 3 is crucial in determining the effector lineages of helper T cells via inducing distinct transcription factors in the activated T cells. In other words, distinct cytokine(s) stimulation is required for the commitment of T cells to each helper T cell lineage. For instance, IL-12 and IL-4 induce the polarization of Th1 and Th2 cells, respectively. IL-6 together with TGF-β drives Th17 cell polarization, while IL-6, IL-12, IL-21 or IL-27 has been shown to promote the polarization of follicular helper T cells (2).
Upon stimulation by infectious agents, allergens, or endogenous inflammatory signals, DCs produce various types of cytokines such as IL-1, IL-6, IL-10, IL-12, IL-23, IL-27, and TNFα that function as signal 3 (3). However, IL-4, a key signal 3 for Th2 polarization, is not produced by DCs. Nevertheless, a series of studies have reported that DCs are required for optimal Th2 responses in vivo, indicating the requirement of DCs in Th2 differentiation. This review focuses on our current understanding of how DCs affect Th2 responses despite their inability to produce IL-4.
DENDRITIC CELL REGULATION OF Th1, Th17, AND REGULATORY T CELLS
Th1 cells
IFNγ-producing Th1 cells are generally induced in response to viruses, intracellular pathogens, and protozoa. Th1 cells activate macrophages, NK cells, and CD8+ cytotoxic T cells to efficiently clear pathogens and pathogen-infected cells. These pathogens are first recognized by pattern recognition receptors expressed on innate immune cells, including DCs. TLRs expressed on the surface or endogenous compartment are important in priming DCs to induce Th1 differentiation. Surface-expressed TLR4 and TLR5 recognize the pathogen-derived molecules LPS and flagellin, respectively. TLRs in the endogenous compartment include TLR3, which recognizes double-stranded RNA, TLR7 and TLR8, which recognize single-stranded RNA, and TLR9, which recognizes unmethylated CpG DNA. TLR signaling eventually triggers the production of IL-12p70 from DCs, which functions as a key factor for Th1 differentiation (4). However, TLR signaling is known to promote the induction of Th1 or Th2 differentiation in a context-dependent manner. The type of microorganism from which LPS is derived, as well as the dose of LPS, seems to determine whether DCs induce Th1 or Th2 responses. Escherichia coli LPS strongly induces Th1 responses through the production of IL-12p70 from DCs, whereas Porphyromonas gingivalis LPS was found to induce Th2 differentiation (5). It has also been proposed that CD4+ cells receiving low-affinity TcR signals differentiate into Th2 cells, whereas high-affinity TcR signals trigger Th1 responses (6). However, the specific identity of the Th1-priming DC subset is still under discussion. Some studies have reported that CD103+ CD207+ DCs are required for Th1 induction (7), while others have claimed that Th1 priming can occur in the absence of that DC subset under certain immunization conditions (8).
Th17 cells
Th17 cells play a protective role against extracellular pathogen and fungi, and also play a pathogenic role in various autoimmune diseases. IL-23 was initially identified as an important cytokine for Th17 differentiation. IL-23 is composed of an IL-12p40 subunit and an IL-12p19 subunit; therefore, it shares its IL-12p40 subunit with IL-12p70. TGF-β and IL-6 were later revealed to be necessary and sufficient for Th17 differentiation. A recent study suggested that an IRF4-dependent CD11b+ CD103+ DC subset drives Th17 differentiation in mice. CD1c+ DCs are known to be a human counterpart of the CD11b+ CD103+ DC subset in mice, and these DCs also express IRF4, secrete IL-23, and promote Th17 responses (910). In addition to murine CD11b+ CD103+ DCs, Langerhans cells were also shown to be necessary and sufficient for Th17 polarization in the Candida albicans skin infection model (11).
Inflammasome activation in DCs is important for Th17 cell differentiation, since IL-1β promotes Th17 cell differentiation. In the experimental autoimmune encephalomyelitis (EAE) model, heat-killed Mycobacterium tuberculosis used for inducing EAE in Freund's complete adjuvant activates the inflammasome and caspase-1 in DCs, leading to the production of IL-1β. Similarly, hyperlipidemic mice exhibited enhanced circulating IL-17, probably due to increased IL-1β and IL-6 from DCs and macrophages via inflammasome-dependent and independent pathways (1213). In addition, the transfer of autoantigen-pulsed DCs caused EAE in naïve recipient mice, indicating the role of DCs in inducing autoimmune Th17 cells (14).
Regulatory T cells
Foxp3-expressing Treg cells are crucial for preventing auto-immunity by inhibiting immune responses against self-antigens. Treg cells also function to suppress effector T cell responses against pathogens to prevent harmful infection-induced immunopathology such as excessive immune reactions. For this reason, some microorganisms have evolved to prime DCs to induce Treg cells. For instance, C. albicans triggers different types of intracellular signals in DCs to modulate distinct T helper responses depending on their fungal morphotypes. DCs induce Th2/Th17 responses to yeast and Th1/Treg differentiation to hyphae (15). In addition, Bordetella pertussis induces DCs to produce IL-10, leading to the generation of IL-10+ Treg cells (Tr1) (16). Zymosan and LcrV from Yersinia pestis activate DCs through TLR2 and/or TLR6 and this signaling pathway induces Treg cells (1718).
Soluble factors of DCs reported to induce Foxp3+ Treg are TGF-β and retinoic acids (19). DCs convert the inactive form of pro-TGF-β to active TGF-β via integrin αvβ8 on their cell surface. Active TGF-β functions as signal 3 for the polarization of peripheral Treg cells and Th17 cells in the absence or presence of an IL-6 signal, respectively (20). In contrast to Th1, Th2, and Th17 differentiation by DCs, the induction of Treg cells does not require mature DCs expressing high levels of MHC class II and costimulatory molecules and can be induced by immature or partially mature DCs. T cells activated by immature DCs express the coinhibitory molecules CTLA-4 and PD-1 (21). It was reported that CD103+ DCs in the gut induce the differentiation of peripheral Treg cells by providing retinoic acid that drives the Treg cell lineage program (1922).
ROLE OF DENDRITIC CELLS IN Th2 CELL IMMUNITY
Evidences that DCs are necessary for Th2 cell responses
While the role of DCs in inducing Th1, Th17, and Treg cell responses via signal 3 is well established, whether DCs have a similar role in inducing Th2 cells has remained relatively unclear. Nevertheless, a series of studies has suggested that DCs are required for optimal Th2 cell responses in vivo. A study using CD11c-diphtheria toxin receptor (DTR) transgenic mice showed that the depletion of CD11c+ DCs in the lung abrogated Th2 cell-mediated asthmatic features such as eosinophilia, goblet cell hyperplasia, and airway hyper-responsiveness (2324). Diphtheria toxin can deplete both CD11c+ DCs and alveolar macrophages in CD11c-DTR mice. The adoptive transfer of CD11c+ DCs, but not macrophages, restored Th2-type asthmatic symptoms in CD11c+ DC-depleted mice. In a model of infection with the parasitic helminth Schistosoma mansoni, CD11c+ DC depletion caused severely impaired Th2 responses and enhanced IFNγ production from CD4+ T cells (25). Similar diminished Th2 responses in the gut were observed with chronic Heligmosomoides polygyrus infection combined with CD11c+ DC depletion in mice (26).
DCs in mouse spleens can be categorized into CD8α+ and CD8α- DCs. When adoptively transferred, CD8α+ DCs induce Th1 cell responses, while CD8α- DCs lead to Th2 cell responses. The induction of Th1-type responses by CD8α+ DCs is due to the production of a large amount of IL-12p70 by CD8α+ DCs. In contrast, CD8α- DCs induce T cells to produce the type 2 cytokines IL-4 and IL-10, although the exact mechanism by which Th2 cell responses can be enhanced by CD8α- DCs is not clear (2728).
In a mouse model of house dust mite-induced asthma, both CD11b+ conventional DCs (cDCs) and CD11b+ monocyte-derived DCs (moDCs) are key factors in initiating and maintaining type 2 immune responses. CD11b+ cDCs migrate to the lung draining lymph node (LN) and mediate Th2 differentiation, while CD11b+ moDCs produce chemokines that attract granulocytes to the lung (29). In addition, in a mouse model of atopic dermatitis, CCL17+ CD11b+ DCs have been shown to mediate Th2 cell responses in a thymic stromal lymphopoietin (TSLP)-dependent manner (30). These studies collectively demonstrate that DCs are necessary for inducing and/or enhancing Th2 cell responses in allergic inflammation and helminthic infection.
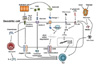
Stimuli that enable DCs to enhance Th2 cell responses
Various types of stimuli can trigger DCs to promote Th2 cell responses (summarized in Fig. 1).
Pathogen-associated molecular patterns (PAMPs)
PAMPs found in microbial pathogens can activate innate immune cells, including DCs, through pattern recognition receptors. Soluble egg antigens from the eggs of the trematode S. mansoni include a glycosylated T2 ribonuclease, termed omega-1, that conditions DCs to induce Th2 cell polarization (31). Omega-1 is one of the PAMPs recognized by the mannose receptor. After being taken up by DCs, omega-1 degrades rRNA and mRNA and thus prevents DCs from synthesizing proteins. This RNase-mediated pathway was found to seemingly program DCs to drive Th2 cell responses (32).
Certain TLRs are known to also induce Th2 cell responses via DCs. For example, Pam-3-Cys, a synthetic TLR2 ligand, conditions DCs to favor Th2 cell responses (33). Pam-3-Cys activates ERK, which subsequently phosphorylates the transcription factor c-Fos, leading to the inhibition of IL-12p70 production and Th1 cell responses. A low dose of LPS was reported to enhance Th2 responses against inhaled antigens through the TLR4-dependent pathway in DCs, whereas a high dose of LPS induced Th1 responses (34). In addition, LPS from distinct pathogens was shown to be capable of conditioning DCs to induce Th2 responses. When LPS from E. coli was used, it induced IL-12p70 production in CD8α+ DCs while LPS from P. gingivalis did not. For T cell responses, E. coli LPS induced Th1 responses with the production of IFNγ, whereas P. gingivalis LPS induced the differentiation of Th2 cells with the production of IL-5, IL-10, and IL-13 (5). Dectin-2, a member of the myeloid C-type lectin receptor family, recognizes house dust mite (HDM) glycans and activates DCs to generate Th2 immune responses through the generation of cysteinyl leukotrienes (35).
Danger signals
Danger-associated molecular patterns released upon tissue damage are usually potent Th2 inducers and are linked to tissue repair process. Adjuvant alum induces Th2 responses in a DC-dependent manner. The underlying mechanism is that alum releases uric acid crystals, which act as a danger signal and activate DCs to induce Th2 cell responses (36). High-mobility group nucleosome-binding protein 1 (HMGN1) is another danger signal that alerts the host defense system, including DCs. HMGN1 was recently reported to play an important role in mediating LPS-induced Th2 responses via DC activation (37).
CELLS OTHER THAN DCs THAT CAN PROMOTE Th2 CELL IMMUNITY
Several types of cells other than DCs are known to mediate Th2 cell immunity either by directly interacting with T cells or by inducing Th2 cell-favoring DCs.
Epithelial cells
Epithelial cells are the first barrier of our body against invading microbes or substances and they secrete various cytokines such as TSLP, IL-25 (IL-17E), and IL-33 in response to allergens and infectious agents. TSLP is expressed in the lungs, intestines, and tonsils (38). TSLPR-deficient mice were reported to show impaired type 2 responses (39) and mice overexpressing TSLP were found to show increased asthmatic symptoms (36). TSLP is known to induce OX40 ligand, which signals T cells via OX40 to potentiate Th2 cell polarization (37). TSLP is also reported to induce the release of chemokines from DCs to attract basophils (38). Basophils can cooperate with DCs in mediating Th2 cell responses by producing IL-4.
DCs highly express IL-33R. The intranasal administration of IL-33 results in type 2 responses, including eosinophilia, IgE secretion, and mucus production (40). In addition, IL-33 stimulates basophils to produce IL-4 in the presence of IL-3 (41) and enhances the release of histamine and IL-13 from mast cells (MCs) and basophils through IgE crosslinking or an IgE-independent pathway. This promotes MC- or basophil-driven inflammation and anaphylaxis (42). IL-33 induces the development of type 2 innate lymphoid cells (ILC2s) that can mediate Th2 cell responses (as described in detail below).
IL-25 is expressed in lung epithelial cells. Overexpression in airway epithelial cells or intranasal administration of IL-25 promotes type 2 inflammation in the lung (43). Lung epithelial cell-derived IL-25 activates DCs to upregulate Jagged 1, a Notch ligand, leading to the induction of Th2 cell responses. IL-25 was also shown to directly stimulate CD4+ T cells to commit to the Th2 lineage (43). In contrast, Th1 and Th17 differentiation is inhibited by IL-25. For example, in autoimmune inflammation, Th17 function was shown to be suppressed by IL-25 (44). Th1-driven inflammation in the gut was also found to be inhibited by IL-25 (45).
These observations collectively demonstrate that cytokines derived from epithelial cells favor Th2 cell immune responses in vivo.
Type 2 innate lymphoid cell
Innate lymphoid cells (ILCs) are TcR-negative innate lymphocytes that can produce T cell cytokines. Like their corresponding helper T cells, ILC1, ILC2, and ILC3 cells express T-bet, GATA3, and RORγt and produce Th1 (IFN γ), Th2 (IL-5 and IL-13), and Th17 (IL-17 and IL-22) cytokines, respectively (46). ILC2s are present in the gut and the airway mucosa of mice. In humans, ILC2s are found in both the gut and lungs of fetuses and adults, and in the palatine tonsils and blood of adults (47). ILC2s enhance Th2-like immune responses, including IgE secretion and eosinophilia, in response to IL-25 and IL-33 (48). Accordingly, increased frequencies of ILC2s are found in diverse allergic diseases such as asthma (49), chronic rhinosinusitis (47), and atopic dermatitis (50). Moreover, ILC2s are one of the major cell types that mediate allergic inflammation against protease allergens in the absence of T and B cells (51).
Interestingly, recent studies have shown that ILC2s can interact with T and B cells. In addition, some ILC2s express MHC class II, which allows them to act as APCs for T cell activation in an antigen-specific manner (52). Furthermore, ILC2s enhances immunoglobulin production from B cells in vitro via IL-5 and costimulatory molecules such as ICOS and CD40L (5253). ILCs generally reside in the tissues rather than lymph nodes and it is unclear whether ILCs can capture antigens in the periphery and then prime naïve T cells in the draining lymph nodes. Further studies are needed to dissect the roles of different ILCs as APCs in vivo.
Basophil
Basophils represent a small population in the circulation. Previously, DCs were known to be dominant APCs for Th2 differentiation. However, recent studies by multiple independent researchers have suggested that basophils act as APCs that trigger Th2 cell responses (5455). The DCsrestricted expression of MHC class II was reported to be insufficient to induce Th2 responses against helminth infection. In this case, basophils are the dominant accessory cell population for Th2 cytokine-dependent immunity (55). A study using a CD11c-DTR bone marrow chimera demonstrated that DCs are neither necessary nor sufficient for inducing Th2 responses, while basophils are responsible for presenting antigens to CD4+ T cells and inducing Th2 responses against the protease allergen papain (54). However, in this experimental model, tissue-resident, wild-type, diphtheria toxin-resistant DCs still exist and need to be considered. A subsequent study that carried out a transient systemic depletion of DCs by the injection of diphtheria toxin into CD11c-DTR mice showed that migratory skin-derived dermal DCs are required for Th2 induction in response to papain and cooperate with IL-4+ basophils (56).
Basophils promote Th2 responses by producing IL-4. Three pathways of IL-4 production by basophils have been suggested. First, the binding of allergen-IgE complexes to FcɛRI triggers IL-4 secretion from basophils. Similarly, IL-3 and IL-33 can stimulate basophils to secrete IL-4 (57). Lastly, PAMPs such as peptidoglycan and other TLR ligands are known to directly stimulate basophils to produce IL-4 (57).
Taken together, basophils represent an important link in the development of Th2 cell immunity, in part by acting as Th2-promoting APCs in certain experimental settings. However, the fact that mice depleted of DCs failed to mount Th2 cell responses raises the question of whether basophil-initiated Th2 cell immunity can be achieved in the absence of DCs.
Mast cell
MCs mainly reside in the tissues, especially in those located at internal-external boundaries where pathogens, allergens, and other external agents continuously invade. Such boundaries include the epithelial surface of the skin and the submucosa of the airways and intestine. This specialized distribution allows MCs to participate in the early recognition of pathogens or allergens (5859). MCs are critical mediators of allergic and anaphylactic reactions and drive IgE-mediated hypersensitivity. MCs contribute to host defenses against parasites such as Trichinella spiralis and Nippostrongylus brasiliensis (4158). It is also proposed that they are involved in autoimmune diseases, including arthritis (60), psoriasis (61), and multiple sclerosis (62).
MCs can be stimulated over timescales of seconds, minutes, and hours by various stimuli such as aggregation of surface FcɛRI (the main pathway of MC activation in allergic responses) and the activation patterns of receptors or signaling molecules (59). Once MCs are activated, they secrete a variety of pro-inflammatory mediators, including histamine, serotonin, proteases (tryptase and chymase), and cytokines such as IL-1, -2, -5, -6, -8, -9, and -13, and TNF α (4158). These cytokines result in the induction of leukocyte chemotaxis, smooth muscle contraction in the bronchial (bronchoconstriction) and gastrointestinal (gastrointestinal motility) tracts, increased vascular permeability, and mucus production by goblet cells.
MCs have been shown to directly and indirectly communicate with DCs and T cells (6364). MC-derived products enhance the maturation and function of DCs. Interestingly, MC-derived histamine (IgE-dependent) and/or type 2 cytokines (IgE-independent) such as IL-25, IL-33, and TSLP inhibit IL-12 production by DCs and subsequent Th1 cell polarization (6566). Moreover, MCs release exosomes that harbor antigen-derived peptides, and these antigen-containing exosomes can stimulate the maturation of DCs leading to enhanced antigen presentation to T cells (66). Thus, it seems likely that MCs also affect Th2 cell immunity by modulating the function of DCs.
FEATURES OF Th2-SKEWING DC SUBSETS
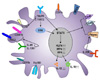
The common features of Th2-inducing DC (Th2 DC) subsets
Unlike other Th cell responses, the mechanisms of Th2 initiation and development associated with Th2 DCs have been poorly understood. Allergic and Th2 immune responses are induced by exposure to allergens, parasites, and proteases (265667). To drive Th2 DCs, Th2-associated cytokines (e.g., TSLP, IL-33, and IL-25) and the expression of FcɛRIII, which has affinity to IgG1 and drives DCs to produce IL-33, are required. DC-produced IL-10 is also needed (3868). Several reports showed that the specific DC subsets are required for facilitating Th2 responses and these specific DC subsets are located in the spleen, lung, and skin. These DC subsets are more efficient in inducing Th2 responses (see above).
Recent studies have defined two surface markers that preferentially expressed on DC subsets specialized in promoting Th2 cell responses. One is macrophage galactosetype C-type lectin 2 (MGL2/CD301b) and the other is programmed death ligand-2 (PDL2) (69). PDL2+ or CD301b+ DCs express Th2 cell-associated molecules, including IL-33R, CD40, CD80, CD86, and OX40L (237071). These DCs also express a specific transcription factor, interferon regulatory factor 4 (IRF4) (9). IRF4 binds to the Il10 gene promoter and induces Th2 cell responses in vivo (72). Mice with Irf4-deficient DCs failed to enhance allergic airway responses, but their impaired Th2 differentiation could be restored by the addition of exogenous IL-10 and IL-33 (7072). In addition, ILC2-derived IL-13 was reported to condition DCs to elicit Th2 responses via the induction of CCL17 (73). TSLP promotes IRF4 expression in DCs through a STAT5-dependent pathway and develops Th2 cell responses through OX40L (7475). However, IRF4 is not sufficient for complete Th2 induction due to its involvement in promoting both Th2 and Th17 responses. Kruppel-like factor 4 (KLF4) facilitates Th2 responses in vivo (76) and acts as a partner molecule to support IRF4 in inducing Th2 cell responses. In addition, CXCR5 is important in Th2 immunity, and H. polygyrus infection-driven Th2 immunity is impaired in mice lacking CXCR5 on DCs or T cells (77). Th2 DCs are not sufficient for the initiation of Th2 responses, although they are necessary for Th2 cell development. With the help of Th2 cytokines and other innate cell populations, Th2 DCs trigger Th2-mediated allergic and inflammatory diseases, including asthma, contact and atopic dermatitis, and delayed-type hypersensitivity. Therefore, targeting Th2 DCs could be a new therapeutic strategy for the treatment of Th2 cell-mediated disease. The proposed phenotypes of Th2 DCs are shown in Fig. 2.
Th2-inducing DC (Th2 DC) subsets
CD301b+ DC
MGL2 (CD301b) is a type II transmembrane lectin (78). CD301b+ DCs are most frequently observed in the dermis and submucosa (7179). CD301b+ DCs were identified as a new type of dermal DCs (dDCs) that polarize Th2 responses and are distinct from migratory Langerhans cells, CD103+ dDCs, CD8α+ DCs, or plasmacytoid DCs in the skin (78). A transcriptomic analysis demonstrated distinct transcript expression profiles between CD301b+ dDCs and CD103+ dDCs, which is a Th1-polarizing DC subset. The CD301b+ DCs expressed lower Th1- and antigen cross-presentation-related molecules than CD103+ DCs did. For example, the expression of the neutrophil attractants Cxcl2 and Cxcl3 was higher, whereas that of Il12b, which is essential for Th1 differentiation, was lower in CD301b+ DCs, compared with the corresponding expression of the same transcripts in CD103+ DCs. CD301b+ DCs also showed lower expression levels of Xcr1, Tlr3, and Clec9a, all of which are involved in antigen cross-presentation. Similar to the gene expression profile, the secretion of IL-12 from CD301b+ DCs was significantly lower than that from CD103+ DCs (78). Additionally, high expression of Th2 cell-promoting molecules such as OX40L, Jagged 1, and IL-33R was observed in CD301b+ DCs (79).
The skin-resident CD301b+ DC subset is a critical initiator of contact hypersensitivity responses (CHR) in vivo (7178). The subcutaneous injection of fluorescein isothiocyanate (FITC)+ CD301b+ dDCs isolated from mice sensitized with FITC (80) to naïve mice sufficiently induced CHR and Th2 immune responses. In contrast, FITC+ CD301b- DCs failed to induce CHR and Th2 humoral responses (7178). In addition, CD301b+ DC-induced Th2 responses in CHR are dependent on TSLP (30). CD301b+ DC-depleted mice showed an impaired Th2 cell-mediated immune response against subcutaneous immunization with ovalbumin (OVA) plus papain or alum and N. brasiliensis infection accompanied by severely abrogated IL-4 production from T cells (70). Particularly, DCs bearing antigens of N. brasiliensis are CD301b+ IRF4+ and have an elevated expression of OX40L. These parasite-specific CD301b+ IRF4+ DCs promote Th2 polarization in a TSLP- and OX40L-independent manner (69). BM-derived CD301b+ DCs promote Th2 responses in effector or memory CD4+ T cells but cannot polarize naïve CD4+ T cells into Th2 cells. (70). For the induction of Th2 cell responses, this DC subset requires maturation by CD301b-targeting antigens and additional help from other cells involved in Th2 immunity (e.g., basophils). According to current reports, there are two main mechanisms for promoting Th2 responses by CD301b+ DCs. The first mechanism involves the upregulation of Th2-associated molecules on CD301b+ DCs, for example, IL-33R and CD40 (7179). The other mechanism involves the provision of signals by CD301b+ DC that regulate LNs to maintain an optimal environment for the Th2 cell differentiation of antigen-specific CD4+ T cells (70).
Though mice encode both Clec10a and Mgl2, which are translated into CD301a and CD301b, respectively, humans do not have a gene that encodes CD301b (81). Therefore, further studies need to be performed to identify the human counterparts of mouse CD301b+ DCs.
PDL2+ DC
PDL2 is the ligand for PD-1, a negative regulator of CD4+ T cells, and type I transmembrane glycoprotein (82). PDL2 is expressed in the lung, liver, spleen, LNs, and even thymus. It is also expressed in macrophages and DCs (8283). PDL2 expression on DCs, either those in the lung or those derived from the bone marrow, is strongly induced by anti-CD40 antibody, GM-CSF, IL-4, and IL-13, and inhibited by IFNγ together with LPS, IL-12, and TGF-β (84858687).
PDL2+ DCs include both dermal DCs and Langerhans cells, and can be divided into CD301b- and CD301b+ subsets. PDL2+ DCs are enriched in the skin- and intestine-draining LNs. Skin draining LN-resident PDL2+ DCs exhibit high levels of CD80, CD86, and CD40 (88). Like CD301b+ DCs, PDL2+ DCs can also enhance the Th2 responses of effector and memory T cells but cannot induce Th2 differentiation (70).
The function of PDL2+ DCs in pulmonary allergic inflammation is well characterized in mouse models. In an OVA-alum-sensitized airway inflammation model, increased expression of PDL2 on lung DCs was observed (23). In addition, the intra-tracheal injection of HDM increased the abundance of HDM-bearing PDL2+ DCs in lung (86). Along with these observations in mice, the expression level of PDL2 in biopsies from human asthmatics correlated with the severity of asthma (86). Besides, exposure to aerosol cigarette smoke combined with OVA increased the expression of MHC class II, CD86, and PDL2 on airway DCs. Similar to CD301b+ DCs, PDL2+ DCs do not require OX40L for the induction of Th2 responses. Interestingly, PDL2 expression on pulmonary DCs was reported to be regulated by Th2 cells (84). Additionally, anti-PDL2 antibody treatment in mice challenged with HDM extract suppressed airway hyperresponsiveness while enhancing the production of IgG2a and IL-12p40, both of which are associated with Th1 cell-mediated immunity (86).
CD11b+ DCs
CD11b+ cDCs and moDCs are associated with the development of Th2 responses (78). Upon activation, CD11b+ DCs migrate to LNs by upregulating CCR7 and CD47 expression and induce Th2 polarization (8990). CD11bhi DCs are partly positive for the Th2 DC markers CD301b and PDL2. In particular, CD11b+ CD301b+ DCs were characterized as a crucial DC subset for inducing Th2 responses in a dibutyl phthalate-FITC-induced mouse model of hypersensitivity (717879).
In an HDM-induced asthma model, CD11b+ DCs could sufficiently induce Th2 responses in lung by producing CCL17 and CCL22 (91). The CCL17+ CD11bhi DC subset is dedicated to Th2-mediated skin inflammation by stimulating Th2 differentiation in skin-draining LNs. CD11b+ DCs have also been identified in the Peyer's patches in the intestine and can drive the differentiation of Th2 cells in response to bacteria (92). Th2-mediated responses by CD11b+ DCs are due to epithelial cell-produced TSLP in part. In this regard, TSLPR could be a marker of Th2 DCs (9394). Furthermore, both GM-CSF and IL-4 could promote the expression of IRF4 in mouse CD11b+ DCs and human DCs (95). Although the CD11b+ DC subset is considered to be one of the Th2 DCs, CD11b+ DCs could not be used as a target for the treatment of Th2-related diseases, since this molecule is also expressed on other types of innate immune cells.
Concluding Remarks
Since aberrant Th2 cell responses to environmental antigens causes diverse immune disorders in mucosal tissues and in the skin, it is important to understand the polarization and maintenance of Th2 cells. Although the source of IL-4 for initial Th2 cell polarization remains unclear, the identification of specialized subsets of dendritic cells that preferentially enhance Th2 cell responses has shed new light on the intricate mechanisms of type 2 immunity and related diseases. Further studies are needed to identify (i) the environmental cues that generate (or differentiate) the Th2-promoting dendritic cells, (ii) the molecular mechanisms by which they mediate Th2 cell responses, and (iii) their role in Th2-related human diseases. The outcomes of such further studies will not only broaden our understanding of Th2 cell responses, but may also facilitate the development of new therapeutic approaches for allergic disorders in humans.
 XML Download
XML Download