

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Ginseng (Panax ginseng) has been widely used in traditional oriental tonic medicine for thousands of years to treat various diseases and to maintain body homeostasis. Many reports show that ginseng has multifunctional biological effects in immune function, anti-inflammatory, anti-cancer, anti-oxidant, metabolic processes (anti-diabetic) and the neuro-endocrine and cardiovascular system (blood pressure regulation) (123456). Ginseng contains many active ingredients including ginsenosides, polysaccharides, peptides, phytosterols, polyacetylenic alcohols and fatty acids (247). Among them, ginsenosides are known to have the most pharmacological and immunological activity (48). In the case of Korean ginseng, 38 ginsenosides have been identified and classified into three groups: protopanaxadiol (PPD), protopanaxatriol (PPT) and oleanane (4).
Recent investigations have demonstrated that ginsenosides are responsible for regulation of the immune response. It has been reported that ginsenoside Rg1 regulates the innate immune response in dendritic cells and macrophages by differentially modulating the production of inflammatory cytokines (910). Rg1 also increases CD4+ T cell activity and modulates Th1/Th2 differentiation in vitro and in vivo (1112). In addition, ginsenoside Rb1, Rd and Re elicit a Th1 and Th2 immune response (13141516), and recent studies have demonstrated that these ginsenosides (Rg1, Rb1, Rd, Re and Rg3) have immunological adjuvant activity to enhance the immune response (17181920212223).
Mature IgM+ B cells undergo Ig class switch recombination (CSR) at the switch region on the heavy chain locus to produce other Ig isotypes (IgG, IgA and IgE) and this class switching is selectively induced by cytokines such as IL-4, IFN-γ and TGF-β1 (24). In addition, expression of germline transcripts (GLTs) for each switch region is a prerequisite for each Ig CSR process (25). That is, the expression of GLα transcripts induces IgA CSR.
As mentioned above, ginsenosides act as adjuvants and then elicit both a humoral antibody response and a T cell mediated immune response. However, the direct effects of ginsenosides on the B cell response have not yet been investigated. To address this, we purified B cells from mouse splenocytes and examined the effects of red ginseng extract (RGE) and ginsenosides on B cell proliferation, antibody production, and expression of GLTs in vitro. Our study reveals that ginsenoside Rg1 and 20(S)-Rg3 selectively induce IgA production and GLTα expression by LPS-activated mouse B cells.
MATERIALS AND METHODS
Animals
BALB/c mice were purchased from Damool Science (Daejeon, Korea) and maintained on an 8:16 h light:dark cycle in an animal environmental control chamber. Eight- to twelve-week-old mice were used, and animal care was in accordance with the institutional guidelines of the Institutional Animal Care and Use Committee of Konyang University.
Purification of B cells, cell culture, and reagents
Mouse splenic B cells were purified by positive selection of B220+ cells using anti-B220 microbeads or by depletion of CD43+ cells using anti-CD43 microbeads and high-gradient magnetic cell separation (MACS; Miltenyi Biotec, Bergisch Gladbach, Germany) according to manufacturer's instructions. Briefly, BALB/c mouse spleen cell suspensions were washed with HBSS (WelGENE, Daegu, Korea) and treated with 0.83% ammonium chloride to lyse the red blood cells. Spleen cells were treated with either anti-mouse B220 microbeads or anti-mouse CD43 microbeads and separated using a LS column and MACS Separator (Miltenyi Biotec, Auburn, CA, USA). The purity of B cells (≥98%) was assessed by FACSCalibur (BD Biosciences, San Jose, CA, USA) after staining the cells with anti-CD43 FITC (eBioscience, San Diego, CA, USA) and/or anti-B220 PE (BD Biosciences). Cells were cultured at 37℃ in a humidified CO2 incubator (Forma Scientific, Marietta, OH, USA) in RPMI-1640 medium (WelGENE) supplemented with 10% fetal bovine serum (PAA Laboratories, Etobicoke, ON, Canada). Purified B cells were stimulated with LPS (1 µg/ml, InvivoGen, San Diego, CA, USA; 12.5 µg/ml, Sigma-Aldrich, St Louis, MO, USA), red ginseng extract (200 µg/ml, Prepared by Dr. JE Choi, Chungnam National University, Daejeon, Korea) (26), ginsenoside Rb1, Rg1, 20(S)-Rg3 (each 2 µg/ml, Ambo Institute, Daejeon, Korea) and TGF-β1 (0.2 ng/ml, R&D Systems, Minneapolis, MN, USA).
Cell proliferation assay
Cell proliferation was determined using an EZ-Cytox cell viability assay kit (Daeil Lab Service Co, Seoul, Korea) as previously described (27). Briefly, 20 µl of EZ-Cytox kit reagent was added to each well of a 96-well microplate and then incubated at 37℃ in a humidified CO2 incubator for 3 h. After incubation, the optical density (OD) was measured at a wavelength of 450 nm using an Absorbance Microplate Reader (BioTek Instruments, Inc., Winooski, VT, USA).
Isotype-specific ELISA
Antibodies produced in B cell cultures were detected using isotype-specific ELISA. Affinity-purified anti-isotype specific antibodies were added at 1.2 µg/ml in 0.05 M sodium bicarbonate buffer (pH 9.5) to 96-well U bottomed polyvinyl microplates (Falcon, Becton Dickinson & Co., Oxnard, CA, USA). Plates were washed with PBS containing 0.05% Tween-20 (PBST) followed by overnight incubation at 4℃, and blocked for 1 h with 0.25% BSA solution. After washing, 50 µl of standard myeloma proteins and culture supernatants were added to each well and incubated for 1 h at 37℃. After washing, horseradish-peroxidase (HRPO) conjugated anti-isotype specific antibodies (Southern Biotechnology, Birmingham, AL, USA) were added to each well and incubated for 1 h. Plates were then washed, and TMB substrate (BD Biosciences) was added. After incubation, 0.05 M sulfuric acid was added to each well, and colorimetric reaction was measured at 450 nm with an Absorbance Microplate Reader.
RT-PCR
RNA preparation, reverse transcription, and PCR were performed as previously described (28). The following PCR primers were synthesized by Bioneer (Daejeon, Korea): GLTα, forward 5'-CTACCATAGGGAAGATAGCCT-3', reverse 5'-TAATCGTGAATCAGGCAG-3' (product size, 206 bp); GLTγ1, forward 5'-CAGCCTGGTGTCAACTAG-3', reverse 5'-CTGTACATATGCAAGGCT-3' (product size, 532 bp); GLTγ2b, forward 5'-GGGAGAGCACTGGGCCTT-3', reverse 5'-AGTCACTGACTCAGGGAA-3' (product size, 318 bp); β-actin, forward 5'-CATGTTTGAGACCTTCAACACCCC-3', reverse 5'-GCCATCTCCTGCTCGAAGTCTAG-3' (product size, 320 bp). All reagents for RT-PCR were purchased from iNtRON Biotechnology (Seongnam, Korea). PCR for β-actin was performed in parallel to normalize the cDNA concentrations within each set of samples. Aliquots of the PCR products were resolved by electrophoresis on 2% agarose gels.
RESULTS AND DISCUSSION
Effects of red ginseng extracts on mouse B cell proliferation and antibody production
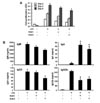
Red ginseng is usually produced by steaming ginseng root at 95~100℃ for 2~4 h, and then drying at 50~60℃ until the moisture content is <15% (29). In this study, we used two kinds of Korean red ginseng extracts (RGEs), which were prepared by water extraction (26): RGE1 was extracted at 75℃ for 24 h; RGE2 was extracted at 85℃ for 12 h. We first examined the effect of RGEs on B cell proliferation in vitro. As shown in Fig. 1A, both RGE1 and RGE2 substantially inhibited LPS-induced mouse splenic B cell proliferation. Next, we determined the effect of the RGEs on antibody production by B cells. The RGEs induced IgA production while decreasing other isotypes including IgM, IgG1 and IgG2b (Fig. 1B). RGE alone did not stimulate B cell proliferation or antibody production (data not shown). Thus, the RGEs selectively induced IgA production by LPS-stimulated mouse B cells. Further studies will be required to analyze the ginsenoside composition of the RGEs and to verify which ginsenoside(s) contributes to the selective induction of IgA production.
Ginsenoside Rg1 and 20(S)-Rg3 increase IgA production and induce expression of germline α transcripts by LPS-stimulated mouse B cells
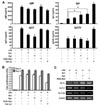
The most abundant ginsenosides in Korean red ginseng are Rb1 and Rg1 (4). In addition, Rg3 [20(R)-Rg3 and 20(S)-Rg3] is one of the most effective ginsenosides for cancer, inflammation, and obesity, and 20(S)-Rg3 is found only in red ginseng. It has been reported that ginsenoside Rb1, Rg1, and Rg3 regulate the T cell-mediated immune response (11121323). Therefore, we first examined whether Rb1, Rg1 and 20(S)-Rg3 have a direct effect on antibody production by mouse B cells. None of the ginsenosides alone were able to stimulate B cell proliferation or antibody production (data not shown). As shown in Fig. 2A, ginsenoside Rg1 and 20(S)-Rg3 significantly increased IgA production (1.7- and 1.9-fold increase, respectively, p <0.05) by LPS-stimulated B cells, while Rb1 had no significant effect on IgA production. Rg1, 20(S)-Rg3 and Rb1 did not increase IgM, IgG1 and IgG2b production. Rather, IgG1 production was decreased by Rb1 and Rg1. TGF-β1 is a well-known IgA and IgG2b class-switching factor (3031). We observed that TGF-β1 selectively enhances IgA and IgG2b production by LPS-activated B cells (Fig. 2A). All three ginsenosides had no effect on B cell proliferation (Fig. 2B). Furthermore, we observed that Rg1 and 20(S)-Rg3 selectively induce expression of GLTα in LPS-activated B cells (Fig. 2C). Our results show that, at least in part, ginsenoside Rg1 and 20(S)-Rg3 induce GLα transcription, resulting in IgA production by B cells.
In summary, in this report, we show for the first time that RGE and ginsenoside Rg1/(20)-S Rg3 can directly drive B cells to produce IgA. Further investigation is need to explore the mechanism(s) by which the ginsenosides induce GLα transcription. It will also be interesting to examine the direct effects of other ginsenosides, such as Rc, Rd and Rh1, on antibody production because it has been reported that intravenous injection of ginsenoside Rc and Rd differentially regulate antibody production (32) and ginsenoside Rh1 can alleviate inflammatory symptoms in atopic dermatitis via reduction of IgE (33).
Rats fed a ginsenoside diet had a significantly higher number of IgA+ cells in the jejunal lamina propria, suggesting that ginsenosides enhance IgA production and support a protective effect against mucosal infections (34). This, together with our results, suggests that ginsenosides [especially, Rg1 and 20(S)-Rg3] could be a selective adjuvant to enhance IgA production by B cells and could be used as potential agents for a mucosal vaccine.
 XML Download
XML Download