

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Eosinophils are multifunctional leukocytes that have been implicated in the pathogenesis of Th2-type inflammatory processes, including helminth infections and allergic diseases. The cytoplasm of mature eosinophils contains numerous secondary granules such as eosinophil peroxidase, eosinophil cationic protein, eosinophil-derived neurotoxin, and major basic protein, and the exocytotic release of these granule-derived cytotoxic proteins contributes to inflammatory responses induced by eosinophil activation (12). Eosinophils are produced in the bone marrow from pluripotential stem cells, which differentiate into eosinophil progenitors marked by CD34+IL-5Rα+ expression (1). Eosinophil lineage specification is determined by the interplay of several transcription factors, including GATA-1 (a zinc finger family member), PU.1 (an ETS family member), and members of the CCAAT/enhancer-binding protein (C/EBP) family (345). Following differentiation, permissive proliferation and migration of eosinophils from the bone marrow to the circulation are regulated primarily by IL-5. However, eosinophils account for less than 5% peripheral blood leukocytes in normal humans and have a short life span, making it difficult to study the biological properties of circulating eosinophils in vitro (6).
A human eosinophilic cell line, EoL-1, has been considered a useful in vitro model to study human eosinophils (78). EoL-1 cells can be induced to develop into eosinophilic granule-containing cells by chemical stimuli, including dibutyryl cAMP (dbcAMP) and butyric acid (9). There are several markers for mature eosinophils, including CCR3 and IL-5Rα (1011). Although independently performed studies have revealed maturation of EoL-1 cells following stimulation by either dbcAMP or butyric acid, comparative phenotypic analysis of differentiated EoL-1 cells will contribute to suggesting optimal EoL-1 stimulating conditions compatible with experimental purpose. In this study, we found that stimulation with butyric acid was more effective than stimulation with dbcAMP for induction of EoL-1 cell differentiation. However, both butyric acid and dbcAMP were not sufficient for the expression of CCR3 in EoL-1 cells, and we propose subsequent cytokine treatment of chemically stimulated EoL-1 cells.
MATERIALS AND METHODS
Cell culture
EoL-1 cells (DSMZ, Braunschweig, Germany) were maintained in RPMI 1640 medium (Sigma, St. Louis, MO, USA) supplemented with 10% FBS (Gibco Laboratories, Grand Island, NY, USA) in 5% CO2 at 37℃. EoL-1 cells were induced to differentiate by the addition of dbcAMP (Sigma-Aldrich) or butyric acid (Sigma-Aldrich) for 9 days. The cell concentration was adjusted to 5×105/ml every 3 days.
Morphological analysis
For morphological analysis, cultured EoL-1 cells were spun at 500 rpm for 5 min on glass slides (Cytospin 3, Shandon, Pittsburgh, PA, USA). The slides were air-dried, stained with Diff-Quik stain solution (Sysmex, Kobe, Japan) and observed using the CX41 microscope (Olympus, Tokyo, Japan).
Flow cytometry
Aliquots of EoL-1 cells, before and after stimulation with dbcAMP or butyric acid, were resuspended in FACS buffer (PBS containing 10% FBS, 10 mM EDTA, 20 mM HEPES, and 1 mM sodium pyruvate). After Fc receptor blocking with human TruStain FcX (Biolegend, San Diego, CA, USA) for 15 min at 4℃, the cells were stained with antibodies against FITC anti-human CCR3 (R&D, Minneapolis, MN, USA) or PE anti-human IL5Rα (R&D). For analysis of viability, cells were incubated with 7-amino-actinomycin D (Biolegend) for 10 min at room temperature. Each sample was analyzed using FACSCalibur (BD Bioscience, San Diego, CA, USA) and the data were processed with FlowJo software (Tree Star, Ashland, OR, USA).
Real-time PCR
RNA from unstimulated and dbcAMP or butyric acid treated EoL-1 cells was extracted using QIAzol lysis reagent (Qiagen, Hilden, Germany) and was column-purified with an RNeasy Mini Kit (Qiagen). The purified RNA (500 ng) was treated with DNase I (New England Biolabs, Ipswich, MA, USA), and cDNA was synthesized using the iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA). Real-time PCR was performed using iQ SYBR Green Supermix (Bio-Rad). PCR was performed using a CFX96 Real-Time System (Bio-Rad). Sequences of primers are shown in Table S1.
RESULTS AND DISCUSSION
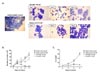
Treatment of EoL-1 cells with either 100 µM dbcAMP or 0.5 µM butyric acid for 6 to 9 days is known to induce eosinophilic maturation (91213). EoL-1 cells represent undifferentiated promyelocytic eosinophils possessing a large nucleus with prominent nucleoli (Fig. 1A) (14). After treatment with dbcAMP or butyric acid, we observed morphological signs of EoL-1 differentiation, including nuclear lobulation and increased proportion of cytoplasm to nucleus (Fig. 1A). However, compared to the group stimulated with 100 µM dbcAMP, EoL-1 cells stimulated with 0.5 µM butyric acid showed remarkable increase in apoptotic populations exhibiting cellular shrinkage and nucleus condensation (Fig. 1A). Consistent with the morphological findings, stimulation with 0.5 µM butyric acid significantly inhibited the proliferation of EoL-1 cells after 2 days (Fig. 1B). The proliferation of EoL-1 cells treated with dbcAMP (100 µM and 10 µM) or 0.05 µM butyric acid increased in a time-dependent manner during 8 days of incubation (Fig. 1B). Additionally, compared to that in other groups, the number of 7-amino-actinomycin D-positive nonviable cells remarkably increased in the group stimulated with 0.5 µM butyric acid (Fig. 1C). These data indicate that stimulation of EoL-1 cells with 0.5 µM butyric acid for prolonged period is unsuitable for EoL-1 cells in vitro assay in terms of cell viability.
Next, we examined the expression of PRG2, EPX, CCR3, IL5RA, and GATA1, markers for mature eosinophils, in EoL-1 cells stimulated with dbcAMP or butyric acid. As shown in Fig. 2A, 0.5 µM butyric acid effectively induced the expression of PRG2, EPX, CCR3, IL5RA, and GATA1 in EoL-1 cells than 100 µM dbcAMP in a time-dependent manner. Additionally, the effect of 100 µM dbcAMP stimulation was limited to the expression of PRG2 and EPX, which encode cytoplasmic granules of eosinophils (Fig. 2B). However, it could be plausible that CCR3 levels could be low even in the 0.5 µM butyric acid-treated group, considering the relatively low fold increase of the transcript in these cells (Fig. 2). Consistent with this idea, mean fluorescence intensity (MFI) of CCR3 expression was not significantly higher in 0.5 µM butyric acid-treated EoL-1 cells than in unstimulated control cells (Fig. 3). Technical problems related to reagents used for flow cytometry analysis were excluded by demonstrating relatively robust expression of CCR3 and IL-5Rα in eosinophils obtained from human peripheral blood (MFI=145±5, MFI=309±11, respectively, Fig. S1). Collectively, these data indicate that compared to dbcAMP treatment, treatment with 0.5 µM butyric acid was more effective in inducing phenotypic maturation of EoL-1 cells, and that the treatment duration should be less than 5 days to preserve the viability of the stimulated cells.
Eosinophil lineage-committed progenitors developed in the bone marrow are identified via surface expression of IL-5Rα and mature into eosinophil precursors containing cytoplasmic granules (15). Eosinophil precursors contain a granule-rich cytoplasm, and their permissive proliferation and differentiation into mature eosinophils is regulated by several cytokines, including IL-5, IL-3, and GM-CSF (16). Recently it was reported that eosinophil lineage-committed progenitors or eosinophil precursors in the mouse bone marrow do not express CCR3 (17). IL-5 primarily induces maturation of eosinophils and stimulates eosinophil migration out of the bone marrow to the circulation mediated by eosinophils expressing CCR3. We have also reported the expression of CCR3 in EoL-1 cells treated with IL-3 and GM-CSF following stimulation with dbcAMP (6). Therefore, we suggest that additional cytokine treatment would be needed to induce differentiation of EoL-1 cells into functionally mature phenotype following chemical stimulation.


 XML Download
XML Download