

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
CD99 is a 32 kDa type-I transmembrane glycoprotein that is exclusively found in primates (1) and is strongly expressed in in cortical thymocytes, bone marrow precursor cells and some plasma cells (23). Engagement of CD99 on these cells has revealed that CD99 signaling is crucial to diverse biological functions in these cells, such as adhesion (456), apoptosis (789), proliferation, and differentiation (1011). Previous studies have uncovered some signaling pathways that operate downstream of CD99 engagement (45678111213141516). Specifically, CD99 engagement is reported to augment activator protein-1 (AP-1) activity through mitogen-activated protein (MAP) kinase pathways in a T-cell line (11) and in human breast cancer cell lines (13).
Basic leucine zipper transcription factor, ATF-like (BATF), is an endogenous repressor of AP-1-dependent transcription (1718). Unlike other AP-1 proteins, BATF lacks a transactivation domain. BATF forms transcriptionally inert complexes with Jun proteins that compete with Jun/Fos complexes for the same DNA-binding sites. As shown by studies that utilized reporter assays, BATF inhibits AP-1-dependent luciferase activity in vitro and in vivo (17181920). Inhibition of AP-1-dependent transcription by the overexpression of BATF effectively blocks cell growth (19202122).
In this study, we examined the signal transduction pathway of CD99 in the human myeloma cell line RPMI8226. Surprisingly, CD99 diminished AP-1 activity by enhancing the expression of BATF. This contrasts with the findings of previous studies that investigated other CD99-expressing cells. Engagement of CD99 also reduced the proliferation of RPMI8226 cells. Overall, our results reveal a novel pathway of CD99-mediated signaling and cellular function.
MATERIALS AND METHODS
Cells and antibodies (Abs)
RPMI8226 and Jurcat cells (American Tissue Culture Collection, Rockville, MD, USA) were cultured in RPMI-1640 media supplemented with 10% (v/v) fetal bovine serum, penicillin G and streptomycin. Anti-CD99 monoclonal Abs (YG32 and DN16-PE) were purchased from Dynona Inc. (Seoul, Republic of Korea). For the engagement of CD99, cells were stimulated with 10 µg/ml YG32 Ab and 17 µg/ml anti-mouse immunoglobulin G (IgG) monovalent F(ab) fragments (Jackson ImmunoResearch, Westgrove, PA, USA) for crosslinking. All anti-MAP kinase antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA).
Reverse transcription PCR and quantitative real-time PCR
Total RNA was extracted from cells using the NucleoSpin kit (Macherey-Nagel, Düren, Germany) and reverse-transcribed using the iScript cDNA synthesis kit (Bio-Rad Laboratories, Hercules, CA, USA). The cDNA was amplified using specific primers for CD99 type-I and -II, as described previously (513). Levels of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA were used to normalize expression. Real-time PCR was performed using an iCycler iQ system with the iQ SYBR Green Supermix (Bio-Rad Laboratories). Fold induction was calculated with the comparative Ct method (23), using the expression of ribosomal protein S18 as the reference. The primers used in real-time PCR analyses are listed in Table S1.
Construction of expression plasmid
The human BATF gene was PCR-amplified from cDNA of RPMI8226 cells using the following primers: 5'-GGC GCTAGCGCCACCATGCCTCACAGCTCCGAC-3' and 5'-GCCCTCGAGTCAGGGCTGGAAGCGC-3'. The amplified products were digested using the NheI and XhoI restriction enzymes, and were ligated into the pCDNA3.1- hygro expression vector (Invitrogen, Carlsbad, CA, USA). The resulting expression construct was sequenced to verify the cDNA sequence vectors were used in the reporter assays.
Transfection and luciferase assay
The luciferase assay was performed using the luciferase assay system and the β-galactosidase assay system (Promega, Madison, WI). RPMI8226 and Jurkat cells were transfected using Lipofectamine LTX (Invitrogen) in accordance with the manufacturer's protocol. A standard transfection reaction used 1 µg of DNA, including AP1-Luc (Clontech, Mountain View, CA), pM1-β-Gal (Roche Diagnostics, Indianapolis, IN) and expression plasmid (pCDNA3.1 or pCDNA3.1-BATF). Luciferase activities were normalized with respect to β-galactosidase activities.
Electroporation of small interfering RNA
BATF-targeting and control siRNAs were purchased from Genolution Pharmaceuticals Inc. (Seoul, Korea) (Table S2). RPMI8226 cells (2.0×106) were electroporated using 1 µM BATF-targeting siRNA or control siRNA using a microporator (Invitrogen, Carlsbad, CA). BATF expression was determined using real-time PCR at 48 hours after electroporation.
Carboxyfluorescein succinimidyl ester (CFSE) labeling
1.0×107 cells were labeled with 0.2 µM CFSE and incubated at 37℃ for 10 minutes. Cold fetal calf serum was added to stop the staining reaction. After 3 days of culture, the fluorescence intensity of CFSE was measured using a FACSCalibur flow cytometer and analyzed using FlowJo software (Ashland, OR, USA).
Apoptosis assays
Apoptosis was analyzed by the apoptosis detection kit (BD Biosciences, San Jose, CA, USA) and tetramethylrhodamine ethyl ester perchlorate (TMRE; Molecular Probes, Eugene, OR, USA) staining. For annexin V staining, cells 1×106 cells were suspended in binding buffer and incubated with Annexin V-FITC and propidium iodide for 15 min at room temperature in the dark. For TMRE staining, 1×106 cells were incubated with 50 nM TMRE for 20 minutes at 37℃. The samples were then measured using a FACSCalibur flow cytometer and analyzed using CellQuest-Pro software (BD Biosciences).
RESULTS
The human myeloma cell line RPMI8226 strongly expresses CD99
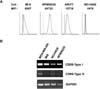
To select a B-cell line for studies of CD99 function, CD99 surface expression was measured in the B-cell lines RPMI8226, ARH77, NCI-H929, and IM-9 by flow cytometry (Fig. 1A). Among the cell lines tested, RPMI8226 cells had the highest surface expression of CD99. In particular, the expression level of CD99 on RPMI8226 cells was higher than on IM-9 cells, in which CD99 engagement triggers intracellular signaling (5). There are two major isoforms of CD99, termed type-I and -II, which have distinct roles in various cell types (513). RT-PCR analysis with primers designed to specifically amplify each isoform showed that only the type-I isoform was detected in RPMI8226 cells, whereas both the type-I and -II isoforms were detected in MDA-MB-435 breast cancer cells, as reported previously (13).
CD99 engagement reduces AP-1 activity and induces BATF expression
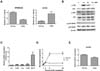
CD99 engagement with the functional anti-CD99 antibody (Ab) is the typical way to trigger CD99-mediated intracellular signaling that culminates in the aggregation, apoptosis, and proliferation of immune cells (456789101124). To elucidate the function of CD99-mediated signaling in RPMI8226 cells, we employed an agonistic anti-CD99 Ab, YG32, to trigger intracellular signaling (1225). We measured changes in AP-1 activity upon CD99 engagement by YG32 in RPMI8226 cells because CD99 has been reported to stimulate AP-1 signaling in other cells (1113). Surprisingly, CD99 engagement reduced AP-1 activity in RPMI8226 cells, whereas it enhanced AP-1 activity in Jurkat cells, as reported previously (Fig. 2A) (11). We next examined MAP kinase activity because MAP kinases are upstream of AP-1, and CD99-mediated signaling activates MAP kinases in Jurkat cells (1112). However, phosphorylation of both extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK) was increased by CD99 engagement in RPMI8226 cells (Fig. 2B). We postulated that AP-1 activity could be regulated by a negative regulator of AP-1 in RPMI8226 cells, rather than by altered MAP kinase signaling mediated by CD99. We measured changes in the expression of AP-1 transcription factors, including BATF, using real-time PCR (Fig. 2C). BATF was identified as being potentially responsible for the reduction in AP-1 levels because it is a negative endogenous regulator of AP-1-mediated transcription. BATF expression was 12-fold higher in CD99-engaged cells than in control cells, whereas the expression levels of Jun, JunB, Fos, and FosB were all less than 2-fold higher in CD99-engaged cells than in control cells (Fig. 2C). Time-course experiments showed that BATF expression was rapidly induced within 8 hours after YG32 Ab treatment, and was sustained at a high level up to 24 hours after CD99 engagement (Fig. 2D). This result is completely different to the effects observed in Jurkat cells, where BATF expression did not increase upon YG32 Ab treatment (Fig. 2E). These results suggest that the induction of BATF expression following CD99 engagement suppresses AP-1 activity in RPMI8226 cells.
BATF regulates AP-1 activity in RPMI8226 cells
To investigate the function of BATF in RPMI8226 cells, we measured AP-1 activity when BATF was ectopically overexpressed. As previously reported in other cells (171819), overexpression of BATF reduced AP-1 activity in RPMI8226 cells, as well as in 293T cells (Fig. 3A). To investigate the extent of BATF involvement in the CD99-induced reduction of AP-1 activity, BATF expression was knocked down by siRNA transfection. Transfection of BATF-targeting siRNA reduced BATF expression by about 50% in comparison to that in cells transfected with non-silencing control siRNA (Fig. 3B). In cells transfected with control siRNA, CD99 engagement reduced AP-1 activity. However, in cells transfected with BATF-targeting siRNA, the CD99-mediated reduction in AP-1 activity was abolished (Fig. 3C). These results demonstrate that induction of BATF expression by CD99 engagement suppresses AP-1 activity in RPMI8226 cells.
CD99 engagement reduces the proliferation of RPMI8226 cells
Given that CD99 engagement suppressed AP-1 activity in RPMI8226 cells, we investigated the effect of CD99 signaling on the proliferation of these cells. Engagement of CD99 by its agonistic Ab YG32 significantly reduced the number of cells recovered after 3 days of culture (Fig. 4A). To investigate whether growth suppression triggered by CD99 engagement was owing to a reduction in proliferation or an increase in apoptosis, apoptosis was evaluated by the TMRE assay and annexin V staining, and proliferation was evaluated by CFSE labeling. After 3 days of culture, the decrease in CFSE fluorescence intensity observed in control cells was evidently retarded in YG32-treated cells (Fig. 4B). In the TMRE assay, there was no marked difference between YG32-treated and control cells (Fig. 4C). Over 24 h of culture, the ratio of annexin V-positive cells to the total number of cells did not significantly differ between YG32-treated and control samples, whereas this ratio significantly increased in bortezomib-treated samples (Fig. 4D). These results suggest that the reduction in the number of cells recovered after CD99 engagement was mostly caused by the suppression of cell division, and not by a marked increase in the rate of apoptosis. We measured the expression of cyclin D1, D2, and D3 using real-time PCR after CD99 engagement because cyclin D proteins are major regulators of proliferation and cyclin D1 is a target gene of active AP-1 (26). The expression levels of cyclin D1 and D3 were significantly lower in CD99-engaged RPMI8226 cells than in untreated cells (Fig. 4E). These results suggest that the decrease in the recovery of RPMI8226 cells following treatment with an anti-CD99 Ab is mainly owing to a reduction in cell proliferation, possibly through reduced AP-1 activity via BATF. Nonetheless, the possible contribution of pro-apoptotic mechanisms cannot be entirely excluded.
DISCUSSION
Variation in the AP-1 response to identical stimuli in different cell types and different microenvironments is well-documented (27). Overall, AP-1 signal transduction is controlled by a variety of factors, including the expression level of individual AP-1 family members, post-transcriptional modification, and the composition of AP-1 molecules on DNA-binding sites of target genes. Inhibition of AP-1 blocks tumor promotion, transformation, progression, and invasion (28). Inhibition of AP-1-dependent transcription by the overexpression of BATF effectively blocks cell growth (19202122). A small number of exogenous stimulating factors, including interleukin-6, leukemia inhibitory factor (29), interferon-β (30), and programmed cell death-1 (31), have been reported to be capable of inducing BATF expression. The present study demonstrated that CD99 is a BATF-inducing factor that reduces AP-1 activity in RPMI8226 cells.
CD99-mediated intracellular signaling involves activation of MAP kinases and increased AP-1 activity in Jurkat acute leukemia cells (11) and breast cancer cells (13). CD99 engagement augmented ERK and JNK activation in RPMI8226 cells. Therefore, the reduction in AP-1 activity by CD99 engagement in RPMI8226 cells prompted us to examine changes in the composition of AP-1 transcription factors following CD99 engagement. In various hematopoietic cells, increased BATF expression inhibits proliferation by reducing AP-1 activity (212231). A reduction in AP-1 activity caused by the c-Jun-dominant negative mutant decreases the expression of D-type cyclins (32). In the present study, CD99 engagement inhibited proliferation and reduced the expression of cyclin D1 and D3 in RPMI8226 cells. Altogether, augmented expression of BATF by CD99 is a plausible mechanism for the inhibition of AP-1 activity, the reduced proliferation, and the downregulation of cyclin D in RPMI8226 cells. Under our experimental conditions, CD99 engagement did not appear to induce extensive apoptosis, in contrast to the findings of previous studies in other cells (789).
The present study showed that CD99 engagement reduces AP-1 activity and the proliferation of RPMI8226 cells. These results suggest a novel pathway of CD99 signaling that controls cell proliferation, and raise the possibility that an anti-CD99 Ab could be used as a novel therapeutic molecule.


 XML Download
XML Download