

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Mesenchymal stem cells (MSCs) are non-hematopoietic adult stem cells typically isolated from adult tissues including bone marrow (BM), adipose tissue, and cord blood (123). Although MSCs are multipotent and can differentiate into various mesenchymal cell types, their immunomodulatory properties distinguish them from other types of stem cells such as hematopoietic stem cells (HSCs), embryonic stem cells (ESCs), and induced pluripotent stem cells (iPSCs). It has become more evident that MSCs possess immunomodulatory properties in vitro and in vivo, as demonstrated by suppression of a broad range of innate and adaptive immune cell types including lymphocytes, natural killer cells, neutrophils, macrophages, and dendritic cells (4). Furthermore, a number of studies have shown that MSCs improve immunological or inflammatory pathologies such as graft-versus-host disease, pancreatitis, and diabetes (567). The immunosuppressive properties of MSCs have already been applied in the development of stem cell therapies for treatment of inflammatory and autoimmune diseases (89). However, the precise molecular mechanisms involved in MSC-mediated immunomodulation remain incompletely understood. Although the cellular and molecular mechanisms of MSCs associated with inflammatory conditions are complex, a variety of soluble factors are critical for MSC-mediated immunomodulation (410). These secreted soluble factors that have been demonstrated to mediate MSC-associated immunosuppression include both protein factors such as transforming growth factor-β (TGF-β), indoleamine 2,3-dioxygenase (IDO), interleukin-10 (IL-10), and hepatocyte growth factor (HGF), and non-protein factors such as prostaglandin E2 (PGE2) and nitric oxide (NO) (11-13). However, blocking any of these soluble factors including TGF-β, IL-10, and HGF failed to achieve a complete ablation of MSC-mediated immunosuppression (14). This functional redundancy suggests that there could be unidentified factors regulating MSC-mediated immunosuppression under different pathological conditions.
Galectins belong to a phylogenetically conserved mammalian family of β-galactoside-binding lectins with a characteristic carbohydrate recognition domain (15). Galectins have pleiotropic functions in the immune system (1617). For example, galectin-9 was first identified as a chemoattractant and activator of eosinophils (1819). Evidence has accumulated to indicate that galectin-9 plays pivotal roles in the modulation of innate and adaptive immunity as well as in a broad spectrum of cellular responses, including cell adhesion, aggregation, apoptosis, and cytokine secretion (20212223). Galectin-9 dampens lymphocytic immune responses by preferentially inducing apoptosis of activated T helper 1 (Th1) and T helper 17 (Th17) cells (2324). In addition, it induces differentiation of naïve T cells into regulatory T cells (Tregs) and concomitantly it suppresses differentiation of T helper 17 cells (Th17) (2526). Furthermore, human galectin-9 has been shown to improve acute graft-versus-host disease in mice (27).
It has become increasingly apparent that the immunomodulatory effects of galectins resemble those of MSCs, suggesting that galectins could be a major MSC-derived factor involved in the suppression of inflammatory responses. This notion is supported by reports that galectin-1 and galectin-3 contribute to MSC-mediated T cell suppression (2829). However, little is known about the functional roles of galectin-9 in the immunosuppression elicited by MSCs. In this study, we examined whether galectin-9 is involved in human MSC-mediated immunosuppression. The functional roles of galectin-9 were evaluated by examining its ability to inhibit mitogen-stimulated lymphocyte proliferation through recombinant protein treatment and knockdown approaches. Our results showed that galectin-9 is induced in human MSCs by pro-inflammatory stimuli and contributes to the suppression of lymphocyte activation mediated by MSCs.
MATERIALS AND METHODS
Isolation and characterization of human MSCs
Bone marrow aspiration was approved by the Inha University Hospital Institutional Review Board (IRB) and written informed consent was obtained from a healthy donor (IRB number #10-51). Highly homogeneous clonal MSCs were isolated from BM as described previously (30). Markers of MSCs were determined by flow cytometry using a number of specific antibodies, including CD14 (BD Biosciences, San Diego, CA, USA), CD29 (Serotech, Kidlington, UK), CD31 (Serotec), CD34 (BD Biosciences), CD44 (Serotec), CD45 (BD Biosciences), CD49f (BD Biosciences), CD73 (BD Biosciences), CD90 (BD Biosciences), CD105 (Serotec), CD106 (BD Biosciences), CD133 (BD Biosciences), CD166(Serotec), HLA-DR (BD Biosciences), HLA-Class I (BD Biosciences), CXCR4 (BD Biosciences), and Oct-4 (Cell Signaling Technology, Danvers, MA, USA). MSCs were positive for CD29, CD44, CD49f, CD73, CD90, CD105, CD166, HLA-Class I, and Oct-4 but were negative for C14, CD31, CD34, CD45, CD106, CD133, CXCR4, and HLA-DR (data not shown). For the assessment of differentiation potential, adipogenic, osteogenic, and chondrogenic differentiations were induced as described (6). MSCs isolated were successfully differentiated into these 3 mesenchymal cell types, indicating the multilineage differentiation potential (data not shown).
Flow cytometric analysis
In order to examine intracellular galectin-9 induction, MSCs were treated with 10 ng/ml TNF-α (BD Bioscience) and 20 ng/ml IFN-γ (BD Bioscience) for the indicated times. After treatment, containing MSCs were washed in phosphate-buffered saline (PBS) containing 0.5% fetal bovine serum (FBS), fixed/permeabilized for 30 min. Then the cells were incubated with fluorescein isothiocyanate (FITC) anti-human galectin-9 antibody (BioLegend, San Diego, CA, USA) for 30 minutes at 4℃ followed by analysis in a flow cytometer (FACSCalibur; BD Biosciences). For analyzing TIM-3-expressing CD3-positive lymphocytes, anti-TIM-3 conjugated with Alexa Fluor 488 (R&D Systems, Minneapolis, MN, USA) and anti-human CD3 conjugated PerCP-eFluor 710 (eBioscience, San Diego, CA, USA) were used.
Enzyme-linked immunosorbent assay (ELISA)
To determine the quality of galectin-9, TNF-a, or IL-17A in the conditioned medium, the wells in a culture plate was coated with 1 µg/ml anti-human galectin-9 antibody (R&D Systems), anti-human TNF-α antibody (BD Biosciences), or anti-human IL-17A antibody (BD Biosciences), respectively, for 24 h at 4℃. After blocking with blocking buffer (1% BSA in PBS), cell culture supernatant was incubated for 24 h at 4℃. After washing, the plate was incubated with a biotinylated anti-galectin-9 (0.2 µg/ml) for 1 h and was incubated with streptavidin-conjugated horseradish peroxidase (HRP) diluted in blocking buffer for 1 h. Then the absorbance at 450 nm of wavelength was measured in a microplate reader (Bio RAD, Foster City, CA, USA).
RNA analysis
To isolate total RNA from MSCs, EasyBlue isolation reagent (Intron Biotechnology, Sungnam, Korea) was used according to manufacturer instruction. cDNAs was synthesized from 1 µg of total RNA by the AccuPower cDNA synthesis kit (Bioneer, Daejeon, Korea). The sequences of the primers used in this study was as follows; galectin-1 (Forward 5'-GGTCTGGTCGCCAGCAACCTGAAT-3' and Reverse 5'-TGAGGCGGTTGGGGAACTTG-3'), gaelcin-9 (Forward 5'-CAGGGCGGTTGGGGAACTTG-3' and Reverse 5'-TGAGGCAGTGAGCTTCACAC-3'), IDO (Forward 5'-GGGCAAATGCTATCATTGGAAAA-3' and Reverse 5'-GGTGGCCGGAGAAGAACATT-3') and GAPDH (Forward 5'-CCACTGGCGTCTTCACCAC-3' and Reverse 5'-CCTGCTTCA CCACCTTCTTG-3'). The PCR products were analyzed by resolution on 1% agarose gel followed by fluorescence image analyzer (LAS4000; Fuji Photo-Film, Tokyo, Japan).
In vitro immunosuppression assay
Thymidine incorporation method was conducted to evaluate lymphocyte proliferation. Peripheral blood mononuclear cells (PBMCs) were collected from healthy donors (IRB #11-104) and separated by ficoll-hypaque density gradient centrifugation. To examine the effects of galectin-9 on the lymphocyte proliferation, 2×105 PBMCs stimulated with 1 µg/ml of phytohemagglutinin (PHA, Sigma-Aldrich, St. Louis, MO, USA) were treated with 1.5 µg/ml of recombinant human galectin-9 (rhGal-9; R&D Systems) for 3 days. In order to determine the immunosuppressive effects of MSCs, 2×105 PBMCs stimulated with 1 µg/ml of PHA were co-cultured with 4×104 MSCs (MSC to PBMC= 1:5) for 3 days. [3H]-thymidine (1 µCi/reaction) was added for the last 12 h of the culture. Lymphocyte proliferation rate was assessed as incorporated radioactivity in counts per minute (cpm).
Carboxy fluorescein succinimidyl ester (CFSE) assay
CFSE assay was conducted to determine whether rhGal-9 or MSCs affect cell division of proliferating lymphocytes. Isolated PBMCs were labelled with 10 µM of CFSE (Invitrogen, Carlsbad, CA, USA) for 10 mins at 37℃. The cell were washed 3 times and re-suspended in the culture medium. The CFSE-labeled cells stimulated with PHA. After 5 days, the cells were harvested and analyzed by flow cytometry.
Galetin-9 knockdown via small interfering RNA (siRNA)
Galectin-9-specific siRNA oligomers (siGal-9; catalog number E-011319-00-0005) and scrambled oligomers (siCon; catalog number D-001950-01-05) were purchased from Dharmacon (GE Healthcare, Lafayette, CO, USA). siRNA transfection was performed according to manufacturer's instruction. Briefly, 5×103 MSCs were seeded onto a well of a 96-well plate. The next day, adherent MSCs were transfected with siGal-9 or siCon. The cells were further stimulated with 10 ng/ml TNF-α and 20 ng/ml INF-γ for 48 h. After knockdown of galectin-9 was confirmed by western blotting, the cells were subjected to in vitro immunosuppression assay and ELISA. For lentiviral shorthairpin RNA (shRNA)-mediated gene knockdown, BLOCK-iT Lentiviral RNAi Expression System was used purchased from Invitrogen. All processes including lentiviral cloning, packaging, production, and infection were performed according to manufacturer's instruction. Target sequences in human galectin-9 (NCBI reference NM_009587.2) are as follows; 5'-GGACTTCAGATCACTGT-3' and 5'-GGAAG ACACACATGCCTTTCC-3'.
Annexin V and propidium iodide (PI) staining
Apoptosis was measured by using an Annexin V/PI detection kit (BD Biosciences) according to manufacturer's instruction. After PBMCs stimulated with PHA were incubated with 1.5 µg/ml of rhGal-9 or were co-cultured with MSCs for 3 days, they were harvested and re-suspended in 500 µl of binding buffer consisting of 0.1 M HEPES/NaOH (pH 7.4), 1.4 M NaCl, and 25 mM CaCl2. Then, the cells were stained with Annexin-V-FITC and PI for 30 min at 4℃. Apoptotic cells were determined by flow cytometry.
RESULTS
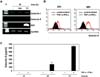
Inflammatory environments or reciprocal interactions between MSCs and activated immune cells are essential for inducing the immunosuppressive effects of MSCs (31). To generate an in vitro condition reflecting the pro-inflammatory environment, we treated human MSCs with the pro-inflammatory cytokines TNF-α and IFN-γ for 24 h. Vehicle-treated MSCs served as a control. We first examined mRNA expression of galectin-9 using RT-PCR. Cytokine treatment strongly induced galectin-9 and IDO in MSCs, while galectin-1 was constitutively expressed regardless of cytokine stimulation (Fig. 1A). Since galectins are expressed in the cytoplasm, flow cytometry was performed after cell permeabilization. Flow cytometric analysis demonstrated significant intracellular expression of galectin-9 in stimulated MSCs within 48 h (Fig. 1B). Galectins can also be secreted in a non-classical pathway bypassing the endoplasmic reticulum and the Golgi apparatus (32). In order to determine whether induced galectin-9 was secreted from MSCs, the culture supernatant was analyzed using ELISA. There was a significant accumulation of galectin-9 secreted from MSCs within 72 h (Fig. 1C). These results demonstrate that in the presence of pro-inflammatory stimuli, galectin-9 expression is highly induced in MSCs and its secretion is stimulated.
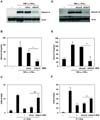
To examine the effects of galectin-9 on activated lymphocyte proliferation, PHA-activated PBMCs were treated with recombinant human galectin-9 (rhGal-9). Treatment with rhGal-9 significantly suppressed lymphocyte proliferation, similar to the inhibition observed for lymphocytes co-cultured with MSCs (Fig. 2A). CFSE assay also revealed that rhGal-9 treatment arrested cell division of activated lymphocytes (Fig. 2B). Thus, the functional effects of galectin-9 and MSCs were consistent, with similar inhibition of lymphocyte proliferation, demonstrating that galectin-9 could be an MSC-derived factor that elicits immunosuppression. Indeed, galectin-9 was secreted in higher amounts in the co-culture of MSCs and PHA-activated PBMCs (Fig. 2C). Release of TNF-α and IL-17A from activated PBMCs was correspondingly decreased by coculture of MSCs (Fig. 2C), suggesting that galectin-9 secreted from MSCs inhibit the production of pro-inflammatory cytokines.
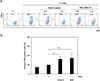
To obtain direct evidence of galectin-9 involvement in MSC-mediated immunosuppression, we examined the effects of galectin-9 knockdown in MSCs on PHA-stimulated lymphocyte proliferation. Galectin-9 was knocked down by siRNA or shRNA-expressing lentivirus. MSCs were transfected with siRNA oligomers. Cells were incubated for 48 h followed by western blot analysis. Significant downregulation of galectin-9 protein expression by galectin-9-specific siRNA oligomers (siGal-9) was observed, while scrambled oligomers (siCon) did not modify galectin-9 protein expression (Fig. 3A). Galectin-9 secretion in the culture supernatant was also decreased as revealed by ELISA (Fig. 3B). Next, we examined the effects of siGal-9-treated MSCs on proliferation of PHA-stimulated lymphocytes. Interestingly, galectin-9 knockdown in MSCs did not suppress lymphocyte proliferation (Fig. 3C). In another experiment, we knocked down galectin-9 using a lentivirus expressing shRNA targeting galectin-9 (shGal-9). After cloning of the lentiviral shGal-9 construct, viral particles were produced in 293FT cells. A lentivirus expressing shLacZ served as a control. MSCs were infected with shGal-9-expressing or shLacZ-expressing lentiviral particles. Dramatic decreases in galectin-9 protein expression and secretion were confirmed by western blot analysis and ELISA, respectively (Fig. 3D). Consistent with the siRNA results, shGal-9-infected MSCs did not suppress proliferation of activated lymphocytes (Fig. 3E). Taken together, these gene knockdown experiments demonstrated that galectin-9 is involved in MSC-mediated immunosuppression. To characterize the the mode of action of galectin-9 in MSC-mediated immunosuppression, we examined whether MSCs or rhGal-9 treatment affect lymphocytes expressing T cell immunoglobulin and mucin-containing domain-3 (TIM-3), a receptor for galectin-9. PHA stimulation induced TIM-3-positive lymphocytes. In the presence of rhGal-9, TIM-3-positive lymphocytes were decreased in a dose-dependent manner (Fig. 4A). Similarly, co-culture of MSCs also reduced TIM-3-expressing lymphocytesin a cell number-dependent manner (Fig. 4A), suggesting that galectin-9 secreted from MSCs binds to TIM-3 to promote galectin-9/TIM-3 internalization. We further examined the effects of galectin-9 or MSCs on apoptosis of activated lymphocytes. In the presence of rhGal-9 or MSCs, cell death of the activated lymphocytes was significantly increased (Fig. 4B). Apoptotic cell death induced by rhGal-9 was comparable to that induced by MSCs, suggesting that galectin-9 contributes to MSC-mediated immunosuppression possibly by inducing cell death of activated lymphocytes.
DISCUSSION
Over the past decade, MSCs have emerged as a promising cell source for the treatment of inflammatory diseases because of their inherent immunomodulatory properties. A prominent feature of MSCs is their capacity to potently suppress T cell activation via multiple mechanisms; they are also able to modulate additional immune cell types including B cells, natural killer cells, dendritic cells, and macrophages (4). A number of studies have demonstrated that MSCs effectively inhibit proliferation, activation, and inflammatory cytokine secretion of activated effector T cells (333435) while they promote the expansion of Tregs (11). Moreover, they have been shown to treat or attenuate several experimental or clinical pathologies such as graftversus-host disease, colitis, pancreatitis, experimental autoimmune encephalomyelitis, and diabetes (36). It is generally accepted that the paracrine mechanism plays a key role in the MSC-induced therapeutic effects in vitro and in vivo, indicating the functional significance of soluble factors released from MSCs (37). Although several soluble factors have been suggested, the redundancy of the immunosuppressive pathways associated with MSCs raises the possibility that uncharacterized soluble factors contribute to the immunomodulation elicited by MSCs.
Although galectin family members have been shown to modulate various lymphocyte functions, including T cell development and activation, apoptosis (17), and cytokine secretion, these proteins have only recently been implicated in MSC-mediated immunomodulation (38). Indeed, there are similarities in the immunomodulatory properties of galectins and MSCs. Galectin-1 is known to be involved in the initiation, amplification, and resolution of inflammation (39). Galectin-1 induces proliferation inhibition, growth arrest, and apoptosis of activated immune cells (4041). Furthermore, galectin-1 suppresses IL-2 production and elevates IL-10 secretion (4243). These functions exerted by galectin-1 have been demonstrated by exogenous addition of a micromolar concentrations of recombinant galectin-1 to T cells. Galectin-3 has pleiotropic functions and can promote apoptosis depending on intracellular or extracellular localization. Extracellular galectin-3 can accelerate activated T cell death whereas intracellular galectin-3 protects cells from apoptosis and promotes T cell proliferation (4445). Exogenous addition of galectin-3 was found to directly initiates apoptosis of thymocytes and T cells, suggesting that it is a potential immunosuppressive factor (46). Conversely, extracellular galectin-3 also promotes pro-inflammatory responses that contribute to the development of autoimmune diseases (47). It has also been shown that galectin-3 induces mast cell degranulation, resulting in the release of pro-inflammatory cytokines and a prolonged inflammatory state (4849). Galectin-9 plays crucial roles in innate and adaptive immunity. In T cell immunity, galectin-9 has been demonstrated to mediate cell death and inhibit the proliferation of activated Th1 and Th17 cells (2450). Galectin-9 signaling has been found to be involved in the development of many inflammatory pathologies, including diabetes, arthritis, graft rejection, and experimental autoimmune encephalomyelitis (25,505152). These immunosuppressive properties, such as potent T cell suppressive activity and tolerance induction make galectin-9 an attractive therapeutic candidate for the treatment of several autoimmune diseases (53).
Galectin-1 and galectin-3 play important roles in MSC-mediated T cell suppression (2829). Although a recent study suggested a functional association between galectin-9 and the immunosuppressive effects of MSCs, the detailed biology of galectin-9 with regard to MSC-mediated immunosuppression has not been investigated. We hypothesized that galectin-9 could be involved in MSC-mediated immunosuppression based on its potent immunoregulatory function. We showed that galectin-9 is significantly upregulated in MSCs and secreted following stimulation with pro-inflammatory cytokines TNF-α and IFN-γ (Fig. 1). The immunosuppressive activity of MSCs is not constitutive but is induced by immune responses (1454). Thus, this inflammation induced factor could be an attractive candidate paracrine regulator. As shown in Fig. 1, galectin-9 expression was not observed in resting MSCs but was strongly induced by treatment with inflammatory cytokines. Through RNA interference-mediated knockdown experiments, we also demonstrated that induction of galectin-9 is required for MSC-mediated lymphocyte suppression (Fig. 3). Based on the comparable induction of lymphocyte apoptosisby galectin-9 and MSCs, we speculate that galectin-9 contribute to MSC-mediated immunosuppression by inducing growth arrest (Fig. 2) and T cell apoptosis (Fig. 4). Galectin-9 is a ligand of TIM-3, a type I membrane glycoprotein that is an inhibitory receptor expressed on terminally differentiated CD4 Th1 cells, Th17 cells, Tregs, natural killer cells, and dendritic cells (24555657). Galectin-9 was previously shown to induce apoptosis of TIM-3-positive Th1 cells through the interaction with TIM-3, indicating that the TIM-3/galectin-9 signaling pathway negatively regulates Th1 immune responses (2458). Our findings also revealed that rhGal-9 treatment or MSCs reduced TIM-3-positive activated lymphocytes (Fig. 4A). Considering that MSCs inhibit T cell proliferation by inducing apoptosis of activated T cells (59), we speculate that galectin-9 secreted from MSCs may could represent a principal MSC-derived factor that binds to TIM-3 and promote the internalization of galectin-9/TIM-3 (60), leading to apoptosis of activated lymphocytes (Fig. 4).
MSCs have been developed as a novel therapeutic option for several inflammatory diseases. Likewise, galectin-9 is considered to have potential to treat pathological immune disorders (6162). Our findings support the notion that galectin-9 contributes to the immunosuppressive function of MSCs. Further investigation is needed to examine how galectin-9 cooperates with other soluble paracrine factors in driving MSC-mediated immunomodulation. Further elucidation of the functional roles and mechanisms of the immunoregulatory factors produced by MSCs could provide foundation for the successful development of stem cell therapies targeting inflammatory conditions and autoimmune diseases.

 XML Download
XML Download