

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
CD4+ T helper (Th) cells are key regulatory immune cells that control a wide range of immune responses against pathogens. Stimulation of the Th cells with Ab against T cell receptor causes activation of Th cells and also induces Th cell proliferation in response to the exogenous IL-2. Activated CD4+ Th cells are subsequently differentiated into several subsets of effector T including Th1, Th2, and Th17, and regulatory T (Treg) cells that are distinguished by signature patterns of cytokine production (1). Th1 cells mainly produce pro-inflammatory cytokine IFN-γ and prevent against tumor development and pathogenic infection, while Th2 cells produce IL-4, IL-5, and IL-13 and mediate non-phagocytic and extracellular pathogen elimination (123). The third effector subset, Th17 cells produce IL-17 and are generated from precursor Th cells by stimulation with TGF-β and IL-6 and induction of RORγt. Treg cells are also generated from naïve CD4+ T cells by the help of TGF-β signaling and FoxP3 induction (45). IL-2 is a major cytokine produced by TCR-triggered Th cells and initiates cell cycle progression and proliferation in an autocrine fashion (6). IL-2 binds to its receptor complex (IL-2R), which is composed of α, β, and γ and activates STAT5-mediated gene transcription through activation of JAK3. This results in accelerated cell proliferation and induction of the IL-2/IL-2R signaling cascade (7). IL-2/IL-2R signaling is also essential for the development of Foxp3 expression in Treg cell development (8).
T-bet is a T-box-containing protein expressed in T cells and is a potent transcription factor that induces Th1 cell development from CD4+ Th precursor cells (9) through direct activation of IFN-γ gene transcription (1011). T-bet deficiency increases the number of Th2 cells at the expense of IFN-γ-producing Th1 cells, resulting in spontaneous asthmatic phenotypes in the lung and acceleration of tumor growth (121314). Restoration of T-bet into T-bet-deficient T cells restored IFN-γ production, attenuating lung inflammation and tumor progression in vitro and in vivo (151617). In addition, T-bet suppresses IL-2 production in developing Th1 cells via direct interaction with NFκB/p65, causing suppression of NFκB/p65 activity on IL-2 gene transcription (18). More interestingly, T-bet modulates the development of RORγt-positive Th17 and FoxP3-positive Treg cells in a RUNX1-mediated manner; additional unknown pathways also contribute (19202122). Finally, T-bet undergoes multiple post-translational protein modifications in Th cells and plays regulatory roles in developing Th cells (2324).
In this study, we investigated whether T-bet has the anti-proliferative activity in different Th cell subsets and whether the anti-proliferative activity of T-bet is dependent on modulation of IL-2 and IFN-γ.
MATERIALS AND METHODS
Mice
Wild type (WT), T-bet knockout (KO), IFN-γ KO, T-bet/IFN-γ double KO, and DTg/KO (TRE-T-bet and rtTA double transgene in T-bet KO background) mice in a C57BL/6 genetic background were housed with free access to water and food. All mice were maintained in specific pathogen-free rooms in an animal facility at Ewha Womans University. All mice handling and experiments were done in accordance with IACUC guidelines at Ewha Womans University (IACUC No. 2012-01-071, 14-030).
In vitro stimulation of CD4+ Th cells
Single cell suspensions were prepared from lymph node and spleen tissues and subjected to isolation of CD4+ Th cells using mouse CD4 micro beads (Miltenyi Biotec, Auburn, CA, USA). Isolated CD4+ Th cells were seeded onto culture plates coated with anti-CD3 Ab in the presence or absence of recombinant human IL-2 (rhIL-2, 10 U/ml). For Th1-skewing conditions, CD4+ Th cells were additionally treated with IL-12 (2 ng/ml) and anti-IL-4 (5 µg/ml). For Th2-skewing conditions, cells were treated with IL-4 (10 ng/ml) and anti-IFN-γ (5 µg/ml). Cells were then cultured for 3 days under Th1- and Th2-skewing conditions and analyzed for cell proliferation activity and cytokine levels. Separately, CD4+ Th cells were isolated from DTg/KO mice and treated with doxycycline to restore the T-bet expression in Th cells, as reported previously (16). Cell supernatants were collected for measuring cytokines, IFN-γ and IL-2 using an ELISA reader (BD Pharmingen, San Diego, CA, USA).
Thymidine incorporation assay
CD4+ Th cells were stimulated with various amounts of anti-CD3 Ab in round-bottomed 96-well plates and then treated with radiolabelled 3H-thymidine (5 mCi/5 ml) to final concentration of 1 µl/well. Cells were harvested 3 days after TCR stimulation and subjected to quantitative analysis. A scintillation beta counter was used to measure radioactivity in DNA recovered from the cells (Microbeta TopCount, Perkin Elmer, Shelton, CT, USA). Three independent experiments were performed for analyzing the results and each experiment was done in triplicate.
Ecdysone-inducible T-bet expression
T-bet cDNA was cloned into the pIND mammalian expression vector. The resulting construct was transfected into human embryonic kidney (HEK) 293 cells (EcR-HEK) that were stably transformed with the regulatory vector, pVgRXR and maintained in the selective medium containing Zeocin (1 mg/ml, Invitrogen, Carlsbad, CA, USA). Empty vector (mock) or the T-bet expression vector was transfected into EcR-HEK cells. G418 (400 µg/ml, Invitrogen) was used to select the following stable cell clones: mock (#1 and #2) and T-bet (#1, #2, #3, and #4). Subcloned cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine, G418, and Zeocin. For induction of T-bet expression, cloned cells were subcultured every 2 days and treated with the Ecdysone analog ponasterone A (PonA, Sigma-Aldrich, St Louis, MO, USA), which was replaced every alternate day.
Luciferase assay
EcR-HEK cells were transfected with mock or T-bet expression vector together with IFN-γ promoter-linked reporter gene and subsequently treated with various concentrations of PonA. Protein extracts were obtained using reporter lysis buffer (Promega, Madison, WI, USA) and used for determining relative luciferase activity using a luciferase assay kit (Promega) and luminometer (Berthold, Bad Wildbad, Germany). Relative luciferase activity was normalized by β-galactosidase activity. The relative activity was expressed as induction fold compared to that of vehicle-treated sample which was set as 1.
RESULTS
Increased proliferation activity in T-bet-deficient Th cells
We examined the proliferation activity of CD4+ Th cells from WT and T-bet KO mice following TCR stimulation. Under non-skewing conditions, CD4+ Th cells proliferated in response to the anti-CD3 stimulus in a dose dependent manner, while T-bet-deficient Th cells showed hyper-proliferative activity in comparison (Fig. 1A). Treatment with excess amount of rhIL-2 had no additional effect on Th cell proliferation in WT or KO mice (Fig. 1B). TCR-induced Th cell proliferation was more prominent in T-bet KO cells regardless of exogenous IL-2 treatment.
Hyper-proliferation of T-bet-null Th cells under Th1- and Th2-skewing conditions
Since T-bet expression is increased by treatment with IL-12 during Th cell differentiation, CD4+ Th cells from WT and T-bet KO mice were stimulated with anti-CD3 Ab and then cultured for 2 days with Th1-skewing cytokines. Like in non-skewing conditions, developing T-bet-null Th1 cells are more proliferative than WT Th cells (Fig. 2A). Exogenous rhIL-2 addition caused no change in cell proliferation of developing Th1 cells (Fig. 2B). Because T-bet deficiency induces Th2 cytokine production, we skewed the cells with IL-4 cytokine to eliminate the developmental difference between WT and T-bet KO Th cells. In response to anti-CD3 stimulation, developing Th2 cells were much more proliferative compared to cells in either non-skewing or Th1-skewing conditions (Fig. 2C). Additionally, T-bet KO Th2 cells are more proliferative than WT cells regardless of IL-2 presence (Fig. 2C and D). These results suggest that T-bet deficiency promotes Th cell proliferation in response to TCR stimulation during Th cell development into both Th1 and Th2 cells.
IFN-γ-independent anti-proliferative activity of T-bet
IFN-γ plays an inhibitory role in regulation of proliferation in immune cells; its production was impaired in T-bet deficient CD4+ Th cells, suggesting that the hyper-proliferative activity of T-bet deficient CD4+ Th cells may be due to diminished production levels of IFN-γ. To assess the IFN-γ dependency in the anti-proliferative activity of T-bet, we examined the effect of T-bet on Th proliferation in an IFN-γ-deficient genetic background. Interestingly, T-bet/IFN-γ double KO mice showed increased proliferation in response to TCR stimulation when compared to IFN-γ single KO mice under both Th1-skewing conditions (Fig. 3A) and Th2-skewing conditions (Fig. 3C). This result was unaffected by exogenous rhIL-2 (Fig. 3B and D). These results imply that the anti-proliferative activity of T-bet is independent of IFN-γ production.
T-bet expression is closely associated with IFN-γ induction and IL-2 suppression in Th cells
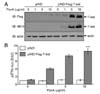
To assess the effect of exogenous T-bet on Th cell proliferation in T-bet KO cells, we generated a Th cell-specific, tetracycline-inducible T-bet transgenic mouse line in a T-bet deficient background (DTg/KO). Treatment of CD4+ Th cells with the tetracycline derivative doxycycline substantially increased T-bet expression in DTg/KO mice (Fig. 4A). Exogenous T-bet expression in T-bet-null cells rescued T-bet function, as assayed by levels of IL-2 and IFN-γ (Fig. 4B). The increased IL-2 expression in T-bet KO cells was suppressed by the induction of T-bet (Fig. 4B). T-bet induction also rescued IFN-γ levels (Fig. 4C). These results confirm that T-bet suppresses IL-2 production in Th cells and that the change in proliferation in T-bet deficient mice is due to loss of T-bet function.
Development of a T-bet expression system in non-T cells
In contrast to IFN-γ, which has no regulatory effect on T-bet-mediated inhibition of Th cell proliferation, IL-2 is a critical factor for regulating Th cell proliferation (18). Additionally, we found that T-bet suppresses IL-2 levels. To rule out a T-bet-mediated effect on IL-2 suppression that influences Th cell proliferation, we established an inducible T-bet expression system in IL-2-independent EcR-HEK cells that were transfected with ecdysone receptor. Subsequent transfection of ecdysone receptor-binding elementlinked T-bet expression vector into EcR-HEK cells induced T-bet expression proportionate to the concentration of PonA (Fig. 5A). IFN-γ promoter activity was increased in a dose dependent manner, mirroring T-bet expression levels (Fig. 5B). Thus, we confirmed functionality of the ecdysone-inducible expression system of T-bet in a non-T cell population that proliferates in an IL-2-independent manner.
Attenuation of cell growth by T-bet induction
To study T-bet's direct role in cell proliferation, EcR-HEK cell clones stably expressing T-bet were selected and cultured for analysis of cell growth rate. Two mock clones and four different T-bet cell clones were treated with PonA. While T-bet expression was not induced in mock clones after treatment with PonA, an increase in T-bet expression was observed to varying degrees in T-bet stable cell clones (Fig. 6A). Cell growth rates of different stable cell clones were calculated by counting total cell numbers following each passage. While PonA had no effect on cell growth in mock clones, cell proliferation rate was substantially decreased by T-bet expression (Fig. 6B). These results suggest that T-bet has an inhibitory role in cytokine independent cell proliferation.
DISCUSSION
This study shows that T-bet induces IFN-γ production and suppresses IL-2 expression in developing Th1 cells. Additionally, the data demonstrate that T-bet-positive Th1 cells are less proliferative than developing Th2 cells. This suggests that T-bet-mediated cytokine regulation may cause delayed cell proliferation during Th1 cell development. However, our results indicate that T-bet plays an inhibitory role in cell proliferation independent of the cytokines IL-2 and IFN-γ. The observed increase in Th cell proliferation caused by T-bet deficiency was also observed in T-bet-deficient IFN-γ KO cells, implying that the anti-proliferative function of T-bet is IFN-γ-independent. In addition, T-bet expression suppressed cell proliferation in non-T cell population whose proliferation is independent of IL-2 and IFN-γ. Collectively, these data show that T-bet has an anti-proliferative activity that is independent of cytokine regulation.
Although it is clear that T-bet suppresses cell growth rate in non-T cell lines and Th cells, the molecular mechanisms remain unclear. Previous research showed that T-bet functions as a transcriptional activator in a direct way (9). Additionally, T-bet undergoes multiple post-translational protein modifications such as phosphorylation at serine, threonine, or tyrosine residues (24) and inhibits gene transcription through interaction with many other transcription factors including GATA-3 (15), NF-κB/p65 (18), NFATp (23), and RUNX1 (19). Thus, expression and post-translational protein modification of T-bet may influence interactions with regulatory proteins involved in cell cycle progression. T-bet may also affect apoptotic cell death, resulting in decreased cell number. The specific effects of T-bet on cell cycle progression and apoptotic cell death remain an interesting area of future research.
IL-2 is a critical factor in early activation of naïve CD4+ Th cells upon TCR stimulation. IL-2 positively regulates its own production through activation of IL-2R signaling and induces cell proliferation (825). As a result, IL-2 influences specification of Th cell development by cooperatively activating several transcription factors and producing signature cytokines (2627). We compensated for the altered levels of IL-2 between WT and T-bet KO cells by treating Th cells with exogenous rhIL-2 and evaluating its effects on Th cell proliferation. Exogenous rhIL-2 addition caused no significant change in proliferation; however, it is possible that endogenous IL-2 production in response to anti-CD3 stimulation could be sufficient to activate Th cell proliferation. In order to confirm whether the anti-proliferation function of T-bet is independent of IL-2 suppression in Th cells, functional assessment of T-bet needs to be performed in the absence of IL-2. However, IL-2 deficiency reduces T cell survival, so it is difficult to identify the specific role of T-bet on Th cell proliferation. Instead, we showed that enforced T-bet expression caused suppression of cell growth rate in non-IL-2-producing cells. How T-bet suppresses cell growth rate in non-IL-2-producing cells is not yet understood. Finally, it still remains unclear whether IFN-γ is induced by T-bet and therefore affects cell proliferation.
T-bet is a critical transcription factor for not only Th1 cell differentiation but also Th2, Th17, or Treg cell development. T-bet undergoes multiple post-translational modification and associates with transcription factor like GATA-3, NFATp, NF-kB, and RUNX1, thus resulting in the suppression of other cell lineage commitment (151824). When compared to the proliferation activity of Th1 and Th2 cells, Th17 and Treg cells are less proliferative. Suppression of Th cell proliferation and suppression of IL-2 production by T-bet may intrinsically affect Th cell differentiation program. It would be interesting to discover the effects and their molecular mechanisms of anti-proliferative activity of T-bet during Th cell lineage commitment.





 XML Download
XML Download