

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Allogeneic hematopoietic stem cell transplantation (allo-HSCT) is an important therapeutic modality used to treat malignancies of hematopoietic origin. Graft-versus-host disease (GVHD) is a major complication following allo-HSCT and triggers non-relapse mortality and substantial morbidity. Gastrointestinal tract damage initiated by preconditioning is a principal pathological cause of mortality from GVHD (1), exposing host epithelial and immune cells to products released by dying (eukaryotic) cells and commensal bacteria (LPS and flagellin) and thereby activating TLRs. TLRs are expressed by various cell types, including innate macrophages, dendritic cells, adaptive T and B cells (2), and non-hematopoietic epithelial cells (3). MyD88 is a primary adaptor that mediates TLR signaling and is known to deliver pro-inflammatory signals (4). However, MyD88 signaling does not always cause inflammation, as demonstrated by the incidence of more severe inflammatory bowel disease in MyD88-deficient mice than that in wild type (WT) mice (5). The recognition of pathogen-derived molecules also occurs through pattern recognition receptors including TLRs, which are widely expressed on parenchymal and nonparenchymal cell types in the liver (6).
Donor bone marrow (BM) cells repopulate the hematopoietic systems of recipients. It was previously reported that MyD88 expression in the allograft non-T cell compartment was important for protection against fatal intestinal GVHD (iGVHD) in two murine models (submitted for publication). To explore the contribution of donor BM MyD88 expression to the development of hepatic GVHD (hGVHD), we induced GVHD using T-cell-depleted (TCD) BM isolated from MyD88-deficient (MyD88KO) donor mice and T cells isolated from WT donors. Here, we observe that the MyD88-dependent expansion of CD11b+ Gr-1+myeloid derived suppressor cells (MDSC) from donor BM aggravated hGVHD severity with increasing T-cell infiltration into the liver, highlighting the importance of enhanced myeloid cell recovery from donor BM for the development of acute hGVHD.
MATERIALS AND METHODS
Mice
Female C57BL/6 (B6, H-2b), B6.Ly-5a (CD45.1+), and B6D2F1 (F1, H-2b/d) mice were purchased from Japan SLC Inc. (Shizuoka, Japan). MyD88KO (B6, H-2b) mice were generated by Kawai et al. (7) and backcrossed for >10 generations onto the C57BL/6J genetic background. All animal experiments were approved by the institutional Animal Care and Use Committees of the Catholic University and Seoul National University.
Induction of acute GVHD
To generate the MHC- and minor histocompatibility antigen-mismatched transplantation model, F1 mice pre-conditioned with total body irradiation (1,100 cGy) were i.v. injected with TCD BM cells (5×106) plus T cells (1×106 or 2×106) purified from allogeneic B6 mice or B6.Ly-5a (CD45.1+) mice, as previously described (8). For LPS pre-treatment of BM donors, LPS (2 mg/kg body weight) from Escherichia coli (O128-B12; Sigma-Aldrich, St Louis, MO, USA) was i.p. injected daily for 3 days.
Cell isolation and flow cytometric analysis
Mononuclear cells were isolated from liver and peripheral lymph nodes (PLN) as previously described (9). MDSC were purified by magnetic-activated cell sorting after incubation with biotin-conjugated anti-Ly6G, according to the manufacturer's instructions (Miltenyi Biotec, Auburn, CA, USA). The following antibodies against mouse antigens were purchased from BD Pharmingen (San Diego, CA, USA): FITC-conjugated anti-CD45.1 and anti-CD8; PE-conjugated anti-CD11b and anti-CD4; PerCP-Cy5.5-conjugated anti-CD8; APC-conjugated anti-Ly-6G (Gr-1); and APC-Cy7-conjugated anti-CD4. PE-conjugated anti-β 2mb Ab (B10.S) was purchased from Santa Cruz Biotechnology (Dallas, TX, USA).
Histologic and confocal microscopic analyses
Formalin-fixed, paraffin-embedded tissue sections were stained with hematoxylin and eosin. Cryosections were incubated with anti-CD45.1-PE and anti-CD107b-FITC (lysosome-associated membrane protein 2, LAMP2), counterstained with DAPI (Sigma-Aldrich) following washing, and examined under a confocal microscope (LSM700; Carl Zeiss, Oberkochen, Germany).
RESULTS
Allogeneic recipients of MyD88-deficient donor TCD BM exhibited less severe hepatic damage despite higher mortality
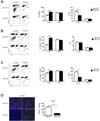
To explore how the absence of MyD88-signaling in donor TCD BM (the non-T cell compartment) affects hGVHD progression, we induced acute GVHD in lethally irradiated F1 mice and then injected T cells purified from WT B6 mice with TCD BM isolated from MyD88KO or WT B6 mice. We have previously demonstrated that allogeneic recipients of MyD88KO TCD BM die of iGVHD earlier than WT B6 TCD BM recipients do (submitted for publication). The GVHD hosts die of severe gastrointestinal damage in this MHC- and minor histocompatibility antigen-mismatched model (1). Unexpectedly, the extent of inflammatory infiltration in the liver was less severe in the MyD88KO TCD BM recipients (Fig. 1A). These findings demonstrated that the lack of MyD88 expression in the non-T cell compartment aggravated the severity of iGVHD in association with serious intestinal damage. In contrast, the presence of MyD88 in the non-T cell compartment exacerbated the severity of hGVHD accompanied by inflammatory infiltration. Consistent with the increased hGVHD-related pathological severity, MyD88KO recipients displayed significantly lower serum ALT levels compared with those of the WT recipients (40.5±3.3 U/L vs. 81.8±10.4 U/L; p<0.01; n=3-6; Fig. 1B).
Donor T cells heavily infiltrated the livers of allogeneic WT TCD BM recipients
In the allogeneic recipients, irrespective of donor BM origin (i.e., MyD88KO or WT), all of the leukocytes infiltrating the GVHD target organs originated from the donor (data not shown). To more clearly compare the extent of tissue infiltration by donor T cells between the two groups of GVHD hosts, T cells isolated from congenic B6.Ly-5a B6 mice (CD45.1+) were used to induce GVHD. The proportion of CD45.1+ T cells in tissue-infiltrating leukocytes was higher in the livers of MyD88KO TCD BM recipients than that of WT TCD BM recipients, because CD45.1- non-T cell population of the former had much less cells than those of the latter (Fig. 2B). However, the absolute numbers of infiltrating CD45.1+ T cells were lower in MyD88KO TCD BM recipient livers than those of WT TCD BM recipient livers, because the total cellularity levels in the liver were lower in the former (Fig. 2A). Additionally, we quantified CD4+ and CD8+ donor T cell subtype proliferation with respect to the presence of MyD88 in this GVHD model. Differences were evident when the levels of CD8+ cell subtypes were compared with those of CD4+ cells (Fig. 2B, C). Moreover, confocal microscopic analyses revealed that CD45.1+ cells were scarcer in the liver of MyD88KO TCD BM recipients than that in WT recipient livers (Fig. 2D).
Reduced expansion of CD11b+Gr-1+ cells in the allogeneic MyD88KO TCD BM recipients
CD45.1- non-T cell compartments, which were notably rare in the MyD88KO TCD BM recipient organs analyzed (Fig. 2A), were comprised primarily of CD11b+Gr-1+ cells (Fig. 3A). The low proportion of CD11b+Gr-1+ cells in the MyD88KO group was attributed to the low levels of expansion from transplanted MyD88KO TCD BM in response to pro-inflammatory stimuli produced in the GVHD hosts. This is because the proportions of CD11b+Gr-1+ cells in naïve BM and in different peripheral organs of MyD88KO mice (i.e., prior to transplantation) were comparable to those of WT B6 mice (data not shown). However, the numbers of LAMP2+ neutrophils did not differ significantly between the two groups (Fig. 3B), indicating intact generation of neutrophils in the MyD88KO TCD BM recipients. Therefore, the reduced number of CD11b+Gr-1+ MDSC from MyD88KO BM was associated with reduced T-cell infiltration into the liver, which suggested the possibility that MDSC could enhance T cell infiltration into the liver (Fig. 2B).
Supplementation of MyD88KO TCD BM recipients with WT MDSC enhanced donor T-cell infiltration into the liver
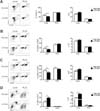
Next, we explored whether supplementation of GVHD hosts with WT MDSC during the course of GVHD could compensate for the lack of MDSC expansion and increase T cell infiltration into the livers of the MyD88KO TCD BM recipients. To this end, MDSC (1×106) purified from WT or MyD88 KO BM were injected on days 3, 5, and 7 post-transplantation into GVHD hosts that had received MyD88KO TCD BM. WT MDSC supplementation increased hepatic T-cell number in the MyD88KO TCD BM recipients. In contrast, enhanced donor T cell infiltration into the liver was not observed following supplementation with MyD88KO MDSC (Fig. 4A). CD4+ and CD8+ T cell numbers were also enhanced in the livers of MyD88KO TCD BM recipients following supplementation with WT MDSC compared with those of animals supplemented with MyD88KO MDSC (Fig. 4B, C). In addition, the number of MDSC increased in the liver following WT MDSC supplementation (Fig 4D). WT MDSC supplementation into the recipients of MyD88KO TCD BM showed more efficient recruitment of donor T cells into livers than MyD88-deficient MDSCs, supporting the idea that MyD88 deficiency in donor BM not only blunted MDSC recruitment or expansion in liver, but also led to generation of MDSC functionally defective in recruiting donor T cells into the liver. These results indicated that functionally intact WT MDSC have a better capability for recruiting or expanding donor T cells than MyD88-deficient MDSC and increase hGVHD incidence.
LPS-injected WT TCD BM recipients displayed high MDSC levels accompanied by a decreased frequency of donor T cells in the liver
To verify the importance of MyD88-mediated signaling by MDSC in the induction of hGVHD, we determined whether transplantation with TCD BM cells containing high levels of MDSC exacerbates hGVHD. As repeated injection of LPS elicits the development of MDSC (10), we injected BM donors with LPS prior to the preparation of TCD BM and confirmed that such pre-treatment increased MDSC frequency and Myd88 expression in the TCD BM inoculums (data not shown). Transplantation with TCD BM isolated from LPS-pre-treated donors significantly decreased the frequency of CD45.1+, CD4+, and CD8+ T cells in the livers compared with that following vehicle-pre-treated TCD BM transplantation (Fig. 5A~C). There was a trend toward an increase in the absolute number of CD45.1+, CD4+, and CD8+ T cells following transplantation with TCD BM injected LPS as compared with that after transplantation with control TCD BM that had been pre-treated with the vehicle even if the statistical significance of this increase was not observed. The proportions and number of MDSC in the livers were higher (Fig. 5D) in the test recipients than those in controls.
DISCUSSION
Following allo-HSCT, donor-derived BM cells repopulate the hematopoietic systems of recipients but may also participate in GVHD pathogenesis, because donor APC crosspresent host antigens to alloreactive donor T cells and exacerbate GVHD (11). However, with the exception of donor APC and T cells, such roles for donor-derived immune cells in hGVHD pathogenesis, have rarely been reported. In the present study, we observed that expression of MyD88 in the non-T cell component of the donor allograft (TCD BM) was important for the induction of hGVHD in a murine model. Lack of MyD88 expression in the TCD BM transplant caused incomplete expansion of MDSC and reduced donor T cell infiltration in allogeneic recipient livers. In addition, an increase in MDSC frequency and MyD88 levels in the TCD BM (achieved via pre-treatment of donors with LPS) enhanced MDSC expansion in the livers.
Inflammation is a key component of a wide range of liver diseases, including hGVHD, steatohepatitis, and acute liver failure due to systemic infections. Of the 10 human TLRs in humans, all are expressed in the liver at the mRNA level, and functional activity of most TLRs was found in the various parenchymal and nonparenchymal liver cell populations (612). It has been demonstrated that expression of the pathogen-sensing machinery in immune cells is important for liver sensitization. In chimera mice that received Propionibacterium acnes or TLR2 and TLR9 ligand treatment to induce liver sensitization, BM-derived immune cells contributed to the damage of the sensitized liver in response to endotoxin (13). In patients with hGVHD, liver biopsies reveal lymphocytic infiltration of small bile ducts with nuclear pleomorphism, epithelial cell dropout, and cholestasis in zone 3 of the liver acinus (14). Our study demonstrated that the extent of inflammatory infiltration was less severe in the MyD88KO TCD BM recipients that had inappropriate BM-derived immune cells, and the appropriate expansion of donor hematopoietic cells might be involved in the hepatic injury observed in hGVHD pathogenesis. A primary effect of MyD88-signaling in hGVHD may be aggravated inflammation via MDSC expansion, whereas the extent of MDSC expansion was reduced in MyD88-deficient mice following cecal ligation and puncture in a model of sepsis (15). In addition, MyD88-deficient mice developed more severe intestinal inflammation than the WT animals in an experimental model of inflammatory bowel disease (5), emphasizing the importance of MyD88-signaling in the protection of the host from the development of several inflammatory diseases involving the intestines.
There is a link between the immunosuppressive properties of MDSC and the extent of GVHD protection following experimental allo-HSCT (1617). MDSC, derived from embryonic stem cells (18) or generated ex vivo by IL-13 treatment (19), reduced the lethality of GVHD. Wang et al. (16), however, observed that injection of immature BM MDSC did not exert such suppressive effects. Therefore, it is probable that both pro-inflammatory and regulatory effects are exerted by MyD88-mediated signaling during the development of GVHD pathophysiology, and that these effects are cell type-specific. Thus, any protective role exerted by MyD88 may be associated with MDSC expansion in the context of GVHD, whereas the pro-inflammatory role may occur primarily in hepatic cells.
In conclusion, our results demonstrate that MyD88-dependent CD11b+Gr-1+ MDSC expansion may be associated with inflammatory cell recruitment and liver injury following transplantation. Selective MyD88 deficiency in hematopoiesis from donor TCD BM could be critical for the early induction of hGVHD following allo-HSCT, providing useful information regarding the development of target organ specificity.



 XML Download
XML Download