

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Commensal microbiota functions not only to serve as targets of host immunity but also as active players in regulation of host physiology and immunity as a result of long-term coevolution of the host and microbes. T cells play central roles in the regulation of anti-microbial immunity and tissue inflammation. Most major T cell types are made in the thymus, although extrathymic generation of some T cell subsets has been described (1,2). T cells are divided into the major TCR-αβ and minor γδ T cell groups. αβ T cells are highly heterogeneous and grouped into CD4+ conventional T cells, CD8+ conventional T cells, NKT cells, and other innate TCRα-expressing T cells such as mucosal-associated invariant T (MAIT) cells (3,4,5,6). CD4+ conventional T cells are further divided into FoxP3+ regulatory and FoxP3- T cells (7,8). FoxP3-CD4+ T cells include various effector and regulatory T cells based on their cytokine phenotype (IFNγ, IL-17, IL-22, IL-4, IL-9, IL-10, IL-35, and/or LAP-TGFβ1) (6,9). These T helper cells include IFNγ+ Th1 cells, IL-17/IL-22+ Th17 cells, IL-4+ Th2 cells, IL-9+ Th9 cells, IL-21+ T-FH cells, and IL-10/IL-35/TGFβ1+ Tregs (9,10,11,12). All of these T helper cell subsets are generated mainly in the periphery from naïve T cells made in the thymus. TCR repertoire and antigen specificity/affinity greatly influence T cell differentiation in the thymus and periphery (13,14). Co-stimulation signals such as CD28, ICOS, CTLA4, OX-40, and PD-1 signaling reciprocally regulate T cell differentiation and effector function (15,16,17). Cytokine milieu during T cell activation is crucial to generate specialized effector versus regulatory T cell subsets (6,9). A mounting body of evidence indicates that nutrients and metabolites provide significant regulatory signals for T cell differentiation (18,19,20,21,22,23). Potentially important roles of gut microbial products such as short-chain fatty acids (SCFAs) have been recently documented (24,25,26). In this review, we will review the recent progress in our understanding of the roles of SCFAs in regulating CD4+ T helper cell differentiation and the impact of this process on tissue inflammation.
ORIGIN, PRODUCTION, TRANSPORT, AND RECEPTORS OF SCFAs
SCFAs refer to free fatty acids containing fewer than 6 carbons and therefore they have short aliphatic carbon-chains. Formic acid (C1), acetic acid (C2), propionic acid (C3), butyric acid (C4), and valeric acid (C5) belong to the SCFA group. These metabolites are distinguished from longer fatty acids such as medium-chain (6-12 carbons) and long-chain free fatty acids. Because they have relatively shorter hydrophobic chains as well as the hydrophilic carboxyl group, SCFAs are water soluble and readily absorbed or transported into cells. SCFAs are produced by gut microbiota as fermentation products, meaning that they are partially oxidized from sugar molecules under anaerobic conditions in the colon. Carbohydrates are good sources of SCFAs but SCFAs can be made from other nutrients such as proteins and peptides albeit at low levels (27). These SCFA precursors, however, are easily degraded by host digestive enzymes in the upper alimentary tract and don't reach the microbiota in the colon in significant amounts for SCFA production. In contrast, digestion-resistant oligosaccharides and fibers (e.g. oligofructose, inulin, pectin, and arabinoxylan) are good sources of SCFAs. Insoluble fibers including cellulose and chitin, however, are not readily fermented by the microbiota and thus do not produce SCFAs at significant levels.
While it is yet to be determined clearly through extensive bacterial isolation and metagenomics studies, available information indicates that bacteria species greatly differ in their genetic make-up of enzymes involved in SCFA production (28,29). Among SCFAs, C2 is relatively more readily produced than C3 and C4 by most enteric and acetogenic bacteria (30). Propionate can be produced by three pathways (i.e. succinate, acrylate, and propanediol) from various sugar molecules such as pentoses, hexoses, and rhamnose (31). Bacteroidetes and some Firmicutes are good producers of C3 mainly through the succinate pathway. Production of C4 requires additional enzymatic processes that extend acetyl-CoA with butyryl-CoA:acetate CoA-transferase, which is active in some bacteria including Roseburia, Eubacterium and Anaerostipes species and Faecalibacterium prausnitzii (27,32).
The combined concentrations of SCFAs produced in the colon reach ~150 mM, making SCFAs the most abundant anions in the colon. SCFAs are absorbed in the colon and either utilized in colonocytes or transported via the portal vein to reach the blood circulation and other organs. The liver and muscle are major systemic organs for SCFA metabolism and consumption. SCFAs enter cells through passive diffusion and carrier-mediated transportation through SMCT1/SLC5a8 and MCT1/SLC16a1 (33,34,35). SMCT1 is a sodium-coupled monocarboxylate transporter 1 for cell intake of SCFAs and related organic acids such as lactate and pyruvate (34). SMCT1 belongs to the SLC5 Na+/glucose cotransporter gene family (33). MCT1 is an H+-coupled transporter for SCFAs and related organic acids and it transports these molecules depending on the net chemical gradients for H+ and monocarboxylates across the membrane (36). Expression of these transporters in the apical membrane of colonocytes, DCs, kidney cells, and/or brain cells has been documented (Table I).
SCFAs activate several G-protein-coupled cell surface receptors (GPCR). GPR41 and GPR43 are major receptors that can be activated by most SCFAs (37). Gut enteroendocrine cells highly express GPR41 and GPR43 (38,39). Other regular enterocytes express these receptors also at functional levels (38,39,40,41). GPR41 is also expressed in adipocytes, renal smooth muscle cells, enteric neuronal cells, and pancreatic cells (Table I) (42,43). The expression of GPR41 is co-regulated with GPR40, a receptor for medium and long-chain fatty acids, because their gene transcription is regulated by the same promoter (44). GPR43 is expressed by granulocytes and some myeloid cells (45,46,47). GPR109a, a receptor for niacin (also called nicotinic acid and vitamin B3), is a receptor also for C4 (48). GPR109a is expressed by gut epithelial cells, adipocytes, macrophages and dendritic cells (Table I). Olfr78 is expressed in the kidney juxtaglomerular apparatus and is activated by C2 and C3 (49). However, T cells do not express these receptors at functionally significant levels (unpublished results) (24). Major cell types expressing these receptors are listed in Table I.
BASIC FUNCTIONS OF SCFAs IN THE BODY
SCFAs, also called volatile fatty acids because of their relatively more volatile nature compared to longer fatty acids, have been studied for more than a century (50,51). These early observations linked SCFAs to diarrhea and ion balance in the intestine. SCFAs are physiologically important in the intestine as they regulate ion absorption and gut motility. Because SCFAs are absorbed first into colonic epithelial cells and can be metabolized in these cells, they profoundly affect the basic biology of intestinal epithelial cells. SCFAs, particularly C4, are used as the major energy source for colonic epithelial cells and regulate their gene expression, proliferation, differentiation, and apoptosis (52). For example, SCFAs promote the production of mucin and gastrointestinal peptide (e.g. LL-37) (53), molecules important for gut barrier function.
SCFAs condition intestinal epithelial cells to make them more readily respond to bacterial products (40). This function is important to prepare epithelial cells for mounting optimal innate immune responses to invading pathogens and commensal bacteria, and therefore helps prevent chronic intestinal inflammatory responses to microbes and their products. In this regard, SCFAs have anti-inflammatory activity in regulating intestinal inflammation (54). Intestinal epithelial cells express GPR41, GPR43, and GPR109a, which mediate a significant portion of the SCFA function (48,55,56,57). These GPCRs activate signaling processes such as RAS, protein kinase A, PI3K, and ERK1/2 for activation of transcription factors such as ATF2 (40,47,48,58,59). Activation of this pathway is important for expression of key immune and inflammatory mediators such as IL-1, IL-6, TNF-α, CXCL1, and CXCL2. Another function of SCFAs is to activate GPR41 and GPR43 on secretory epithelial cells to produce glucagon-like peptide (GLP)-1 (60). Enteric neurons express GPR41 to sense SCFAs for regulation of gut motility (39). To support this, there is a high correlation in expression sites between SCFA receptors and gut endocrine hormones such as GLP-1, PYY, and neurotensin. Another major mechanism for the SCFA regulation of epithelial cells is mediated through inhibition of HDACs by SCFAs (61,62).
Other cell types are also regulated by SCFAs. SCFAs induce the chemotaxis of neutrophils via activation of GPR43 (58,59) and regulate neutrophil degranulation (63,64). SCFAs also regulate macrophages and dendritic cells (DCs) (65,66). SCFAs suppresses NF-kB and the production of inflammatory cytokines such as IL-6 and TNF-α but increases IL-10 secretion from macrophages (67). In contrast, increased C2 levels in alcoholism can increase the expression of inflammatory cytokines in macrophages and even exacerbate the inflammatory response in the liver (68). Thus, the SCFA function in regulation of immune responses may be altered in pathological conditions. SCFAs increase satiety and reciprocally regulate adipogenesis and lipolysis (69,70,71). Adipocytes express GPR41 and are activated by SCFAs to produce leptin (42,43,72). Olfr78 activation promotes renin production from the kidney to regulate blood pressure (49).
COMPLEX ROLES OF SCFAs IN REGULATION OF EFFECTOR VERSUS REGULATORY T CELLS
Early work on C4 revealed its regulatory effect on cytokine production by lymphocytes (73,74). C4 had regulatory effects on production of cytokines such as IL-2, IL-4, IL-5, IL-6, and IL-10 (75). Others observed that C4 induced Fas-upregulation and apoptosis in T cells (76). Smith et al. reported that mice fed with SCFAs had increased numbers of IL-10-producing FoxP3+ T cells in the colon (25). The effect was specific for colonic FoxP3+ T cells, and FoxP3+ T cells in other organs were not expanded after SCFA administration. A mechanism provided by this group for the expanded colonic FoxP3+ T cells was decreased HDAC expression and activity by SCFAs in a GPR43-depednent manner. SCFAs can enter cells through diffusion or carrier-mediated transport and thus do not necessarily go through cell surface receptors. Moreover, T cells do not express GPR43 at significant levels and thus this mechanism remains to be verified. Another group reported that C2 and C3 can directly suppress HDACs and increase histone acetylation at the FoxP3 gene locus for increased transcription (20). Similarly, it was reported that Treg generation was increased by SCFAs as a result of HDAC inhibition by SCFAs and histone H3 acetylation in key regulatory regions of the Foxp3 locus (77). In relation to these reports, inoculation of germ-free mice with SCFA-producing Clostridia groups induced IL-10 and ICOS-expressing FoxP3+ T cells (78). Overall, these studies suggest that SCFAs expand colonic Tregs for immune tolerance.
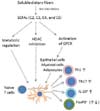
Our group found that SCFAs can increase IL-10, but not necessarily FoxP3, expression in T cells (24). Interestingly, SCFA either positively or negatively regulate induced FoxP3+ cells depending on the strength of T cell activation in vitro. In high T cell activation conditions, SCFAs can even suppress FoxP3+ cell induction promoted by TGFβ1 and T cell activation. In contrast, SCFAs enhance FoxP3+ cell induction at low T cell activation conditions. Independent of FoxP3 regulation, SCFAs increased IL-10 production in all T cell activation conditions (24). We observed that the FoxP3+ T cells even in the colon were not reproducibly regulated by SCFA administration in vivo (unpublished results). These results imply that FoxP3 induction by SCFAs may be regulated by indirect mechanisms through non-T cells. A rather surprising finding was that SCFAs facilitated naïve T cell differentiation into Th1 and Th17 cells in appropriate T cell polarization conditions. Thus, SCFAs can enhance both effector and regulatory T cells depending on the immunological milieu. In support of this, C2 administration via drinking water increased Th1 and Th17 cells in the intestine and secondary lymphoid tissues during C. rodentium infection (40). In the absence of infection, SCFAs increased gut IL-10+ T cells in vivo, which would promote immune tolerance. It appears that SCFAs selectively promote only the right types of T cells required to handle specific immunological conditions (Fig. 1).
POTENTIAL MECHANISMS FOR THE REGULATORY EFFECT OF SCFAs ON T CELLS
Cells can be regulated by SCFAs largely in three different mechanisms (Fig. 1). The first mechanism involves the activation of SCFA-binding G-protein-coupled receptors (GPCRs) such as GPR41, GPR43, GPR109A, and Olfr78. GPCR signaling can regulate cell activation, proliferation, and differentiation. However, T cells do not express any of these receptors at significant levels, according to published information and unpublished microarray data. Thus, SCFA receptors are not likely to be important for direct regulation of T cells by SCFAs. Another pathway is to regulate cell energy status and relevant signaling processes through integration of SCFAs into cellular metabolism. SCFAs can be converted to Acetyl-CoA and integrated into the citric acid cycle (Krebs cycle). Acetyl-CoA is a central molecule that stores energy in the molecule, which is eventually oxidized to CO2 for energy production. As the result, the cellular energy [ATP/ADP] level increases, and this change boosts mTOR activation (79). In T cells, activation of mTOR skews T cell differentiation into effector T cells such as Th1 and Th17 cells at the expense of FoxP3+ T cells (80). mTOR activation also promotes the generation of IL-10+ cells (81). Thus, the SCFA-regulation of cell metabolism and mTOR accounts for the increased generation of Th1, Th17 cells, and IL-10+ cells. The third mechanism is mediated through the HDAC inhibitor activity of SCFAs (Fig. 1). All major SCFAs such as C2, C3, C4 and C5 have HDAC inhibitor activity (24,82,83). Some regarded that C2 does not have the HDAC inhibitor activity but it has clear HDAC inhibitor activity at concentrations (~10 mM) higher than C3 and C4 (~1 mM) (24). Moreover, C2 is maintained at relatively high concentrations in blood (~1 mM). This HDAC inhibitor activity requires the transport of SCFAs into cells and enzymatic inhibition of HDACs. Class I/II HDACs are major targets of SCFA inhibition. While SCFAs do not suppress class III HDAC such as Sirt1, down-regulation of Sirt1 expression by SCFAs was reported (84). Thus, SCFAs may affect a broad range of HDACs for their regulatory effects. Because HDAC inhibition increases the acetylation of histone and other proteins, the impact of this activity is far reaching and affecting a number of genes and proteins. Physical interaction between HDACs and S6K has been reported (85), and S6K is a downstream effector molecule of the mTOR pathway. P70-S6 Kinase 1 (S6K) is hyper-acetylated by SCFAs in T cells, leading to increased mTOR activity in T cells (24).
SCFAs can indirectly affect T cells through their effects on other cells that control T cell differentiation such as DCs (Fig. 1). SCFAs suppress the development of bone marrow progenitors into myeloid DCs in vitro (86). It has been observed that SCFAs also inhibit functional maturation of DCs in vitro (66,86,87,88,89,90). For example, C4 suppressed the maturation of bone marrow-derived DCs and production of IL-12 but increased the expression of IL-23p19 (89). Valproic acid, a branched short-chain fatty acid and potent HDAC inhibitor, suppressed the maturation of human DCs in vitro, inhibiting the up-regulation of T-cell activating molecules such as MHC II, CD80, CD86 and IL-12 (90). While the functional importance is yet to be determined, a report indicates that C4 increased CD1d at the expense of CD1a expression on developing human DCs (88). GPR109a activation affects colonic macrophages and DCs for generation of Tregs and IL-10-producing T cells (91). This effect, however, is not solely due to C4, because GPR109a is a receptor for niacin as well. In this regard, niacin treatment suppressed colitis and colon cancer in a Gpr109a-dependent manner. Moreover, Gpr109a-/- colonic epithelial cells were defective in producing IL-18 in response to C4. More studies are required to separate the niacin from SCFA effect in regulation of GPR109a. Overall, the published results indicate that the regulatory effects of SCFAs have the potential to steer DC development into tolerogenic DCs for promotion of immune tolerance. A caveat is that it remains to be fully determined if SCFAs would exert the same inhibitory effect on DCs in vivo.
REGULATION OF TISSUE INFLAMMATION BY SCFAs
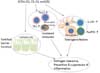
The intestine is the first organ that encounters gut commensal bacteria-derived SCFAs. Therefore, SCFAs have been studied for decades for their effects on inflammatory bowel diseases (IBD). Despite some conflicting reports, high SCFA-producing conditions formed with high levels of dietary fibers are linked to decreased tissue inflammation in the intestine (92,93,94). Oral administration of C4 ameliorated T cell-induced colitis in lymphopenic mice (26). C4 administration attenuated inflammation and mucosal lesions in dextran sodium sulfate (DSS)-induced colitis, an experimental model frequently used for ulcerative colitis (95). However, there is a conflicting report that C4 administration via drinking water worsened the colitis induced by DSS (89). SCFAs also failed to regulate the acute colitis induced with 2,4,6-Trinitrobenzenesulfonic acid (TNBS) (96). These conflicting results may have been obtained due to differences in methods to induce inflammation and regimens to treat the heterogeneous inflammation. To make the function of SCFAs even more complicated, both increased and decreased DSS-induced inflammation in GPR43-deficient mice has been reported (45,97). GPR43-deficient mice had exacerbated inflammation in animal models of colitis, arthritis and asthma (97). GPR43 may modulate gut inflammation, in part, through cytokine production by mononuclear cells (98). GPR43 and GPR41, expressed by tissue cells such as epithelial cells, are also important to prevent chronic inflammation in the intestine following C. rodentium infection (40). Thus, the available information suggests that SCFA receptors play an overall beneficial role in prevention of inflammation (Fig. 2). More work is required to identify the cell types and mechanisms that mediate the beneficial effect of SCFAs in a SCFA receptor-dependent manner.
In humans, C4 enemas had a small ameliorating effect on human colitis patients (99). Treatment of patients with distal ulcerative colitis with C4 (100 mM) was effective in ameliorating disease activity (100). Moreover, treatment of patients with mild to moderate distal ulcerative colitis with combined SCFA enemas (100 mL, twice daily enemas of sodium acetate 80 mM, sodium propionate 30 mM, and sodium butyrate 40 mM) were effective in ameliorating colitis (101). A similar therapeutic effect was observed in ~50% of ulcerative colitis patients who were refractory to a rectal and oral therapy with 5-aminosalicylic acid and corticosteroid (101). SCFAs improved the efficacy of other treatments such as oral mesalazine therapy (102). There is a report that patients with mild to moderate ileocolonic Crohn's disease who were treated with 4 g/day C4 tablets for 8 weeks had decreased clinical activity (103). A caveat is that several large randomized studies found no significant effects of SCFA therapies on ulcerative colitis patients (104,105). These mixed results indicate that SCFAs and their receptors may regulate inflammatory responses only in certain pathological conditions, ameliorating certain types of inflammatory responses while exacerbating other types of responses. Beyond inflammatory bowel diseases, high fiber diets and SCFAs have suppressive effects on respiratory allergic diseases (106). Overall, SCFAs have the potential to work through multiple cell types, including T cells, to exert their regulatory effects on tissue inflammation (Fig. 2).
CONCLUDING REMARKS
The gut microbial metabolites SCFAs profoundly regulate T cell differentiation in the body. Because these metabolites are produced at high levels in the gut, the T cells in the intestine and gut-associated lymphoid tissues are an important cell target for regulation by SCFAs. SCFAs can be transported into the blood and have the potential to regulate T cell activity in systemic tissue sites as well. Beyond T cells, SCFAs regulate the function and phenotype of a number of immunologically important cell types such as epithelial cells, neutrophils, and antigen presenting cells. While the anti-inflammatory activity of SCFAs has been emphasized, SCFAs can also promote the generation of effector T cells and enhance gut barrier function and innate immunity. All of these effects of SCFAs are important to maintain a healthy immune system and to prevent inflammatory diseases. More studies are required to sort out the detailed mechanism of SCFA-mediated regulation of T cells and other immune cells. The current body of literature indicates that SCFAs are not a panacea for inflammatory diseases and may exacerbate certain types of tissue inflammation. Therefore, it is important to identify the types of cells, immune responses, tissue inflammation, and diseases that are highly responsive to SCFA-based therapies.

 XML Download
XML Download