

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
T cells express TCRs for the recognition of peptide epitopes presented on MHC molecules. They acquire diverse TCRs during development via the rearrangement of TCR genes. In the thymus, T cells with TCRs that bind weakly to self-peptides presented on self-MHC molecules survive through a process called positive selection (1). Then, T cells with TCRs that bind strongly to peptide-MHC complexes in the thymus are eliminated through a process called negative selection. Once these selection processes are complete, mature CD4+ or CD8+ T cells are released to peripheral circulation (1).
CD8+ CTLs recognize endogenously processed peptides presented on MHC class I molecules. In particular, CTLs recognize peptides generated via the proteasomal cleavage of viral or tumor antigen proteins and therefore play a critical role in immune responses against virus-infected and tumor cells (2). When CTLs are activated by TCR stimulation, they exert cytotoxic activities through the degranulation of perforin and granzymes and secrete cytokines such as interferon (IFN)-γ and tumor necrosis factor α (2).
After viral infection or vaccination, CTL responses are predominantly directed to a small number of epitopes, although many potential epitopes are present in viral proteins (3). Therefore, there are immunodominant and subdominant epitopes that elicit large and small CTL responses, respectively, in the protein pool of a given virus. This hierarchy of CTL responses is reproducible and predictable in a genetically identical mouse strain (3). The immunodominance of CTL responses is known to be determined by the frequency of naïve CTL precursors with TCRs that recognize a given epitope (4). In fact, a high frequency of naïve CTL precursors often results in immunodominant CTL responses after viral infection (5,6).
Human MHC class I-expressing mice have been established to study human MHC class I-restricted CTL responses in mice. HLA-A2-transgenic mice were developed first and then improved upon because HLA-A2 is the most abundant MHC class I allotype in human beings (7,8,9). Other human MHC class I-transgenic mice have also been developed (10,11,12) and used for several purposes, including the identification of human CTL epitopes and acquisition of human CTL epitope-specific T cells. However, HLA-A2-restricted peptide epitopes were usually poorly immunogenic in HLA-A2-transgenic mice, and their immunogenicity was improved in H-2 class I knockout HLA-A2-transgenic mice (13,14,15).
In the present study, we used HLA-A2-transgenic mice that also express mouse H-2 class I molecules. We found that in many cases, HLA-A2-restricted peptide epitopes were much less immunogenic than H-2 class I-restricted epitopes. We also found that the poor immunogenicity of HLA-A2-restricted epitopes was associated with the paucity of epitope-specific, naïve CTL precursors in HLA-A2-transgenic mice.
Go to : 
MATERIALS AND METHODS
Mice
We used HLA-A2-transgenic mice expressing AAD, which consists of the α1 and α2 domains of the HLA-A2.1 molecule and the α3 domain of the mouse H-2 Dd molecule, under the direction of the HLA-A2.1 promoter (Jackson Laboratory, Bar Harbor, ME, USA) (8,16). These mice expressed AAD in addition to mouse MHC H-2b. Conventional C57BL/6 mice without the HLA-A2 transgene were also used. Mice were kept in accordance with institutional guidelines and used at the age of 6 weeks for peptide immunization.
Peptide immunization
We used ovalbumin (OVA)257 (17), vaccinia virus (VV) A11R198 (18), Japanese encephalitis virus NS4B215 (19), and West Nile virus NS4B215 (19) peptides as H-2 class I-restricted peptide epitopes. We also used influenza A virus (IAV) M158 (20), IAV (PR8) NA231 (21), hepatitis C virus (HCV) core35 (22), HCV core132 (22), HCV (genotype 1b) NS31073 (21), HCV (genotype 1b) NS31406 (23), and Epstein-Barr virus BMLF1280 (20) peptides as HLA-A2-restricted peptide epitopes. Their sequences and MHC restrictions are summarized in Table I. Peptides were purchased from Peptron (Daejeon, Korea) and dissolved in 5% dimethyl sulfoxide phosphate-buffered saline.
Female mice were immunized three times subcutaneously in the hind footpad at 2-week intervals with a mixture of a peptide epitope (100µg), a helper epitope (50µg), lipopolysaccharide (5µg; Invivogen, San Diego, CA, USA), and CpG oligodeoxynucleotide 1826 (5µg; Genotech, Daejeon, Korea). The helper epitope was the H-2 I-Ab-restricted, hepatitis B virus core128 peptide (TPPAYRPPNAPIL) (24). Twelve days after the third immunization, the mice were killed, and splenocytes were prepared for direct ex vivo IFN-γ ELISPOT assays and MHC class I pentamer staining.
rVV infection of mice
We used an rVV expressing HCV (genotype 1b) NS3, NS4A, and part of NS4B (amino acids 1007~1890 of the HCV polyprotein; rVV-HCVNS3-4B) (25). The rVV was derived from the WR strain. Female mice were challenged intraperitoneally with 107 plaque-forming units of rVV-HCVNS3-4B (25). Ten days later, the mice were killed, and splenocytes were prepared for direct ex vivo IFN-γ ELISPOT assays.
Direct ex vivo IFN-γ ELISPOT assays
Direct ex vivo IFN-γ ELISPOT assays were performed as previously described (26). Triplicate cultures of 500,000 splenocytes were established in RPMI-1640 medium containing 10% fetal bovine serum and 2 mM L-glutamine. The splenocytes were stimulated with a peptide epitope (10µg/ml), and 5% dimethyl sulfoxide phosphate-buffered saline was used as a negative control. After 30 h of culture, IFN-γ spots developed. The spots were counted with an ELISPOT reader (CTL, Cleveland, OH, USA), and the number of specific spots was calculated by subtracting the mean number of spots in negative control wells from the mean number of spots in peptide-stimulated wells.
MHC class I pentamer staining
We used PE-conjugated HLA-A0201 pentamers loaded with IAV M158 or HCV core132 peptides (Proimmune, Oxford, UK). After peptide immunization, splenocytes were stained with ethidium monoazide and then with PE-conjugated HLA-A2 pentamer for 20 min at room temperature. Further staining was performed with fluorochrome-conjugated anti-CD3 (BD Biosciences, San Jose, CA, USA) and anti-CD8 (BD Biosciences). The stained cells were analyzed on an LSRII (BD Biosciences) using FACSDiva software (BD Biosciences) and FlowJo software (Tree Star, Ashland, OR, USA).
Enrichment of epitope-specific, naïve CTL precursors
To enrich epitope-specific, naïve CTL precursors from the whole body of a mouse, we harvested the spleen and inguinal, cervical, mesenteric, periaortic, and axillary lymph nodes and prepared a single-cell suspension (27). The cells were stained with PE-conjugated H-2 Kb pentamer loaded with OVA257 peptide or HLA-A0201 pentamers loaded with IAV M158 or HCV core132 peptides (Proimmune) and further labeled with anti-PE magnetic microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany). The microbead-labeled cell suspension was passed through an LS column (Miltenyi Biotec), and MHC class I pentamer-stained cells were enriched. The enriched cells were further stained with fluorochrome-conjugated anti-CD3, anti-CD8, and anti-CD44 (BD Biosciences) and analyzed on the LSRII using FACSDiva and FlowJo software. The frequency of naïve CTL precursors in the mouse was calculated by multiplying the number of cells in the enriched cell population by the percentage of MHC class I pentamer(+) cells (27).
Go to :
RESULTS AND DISCUSSION
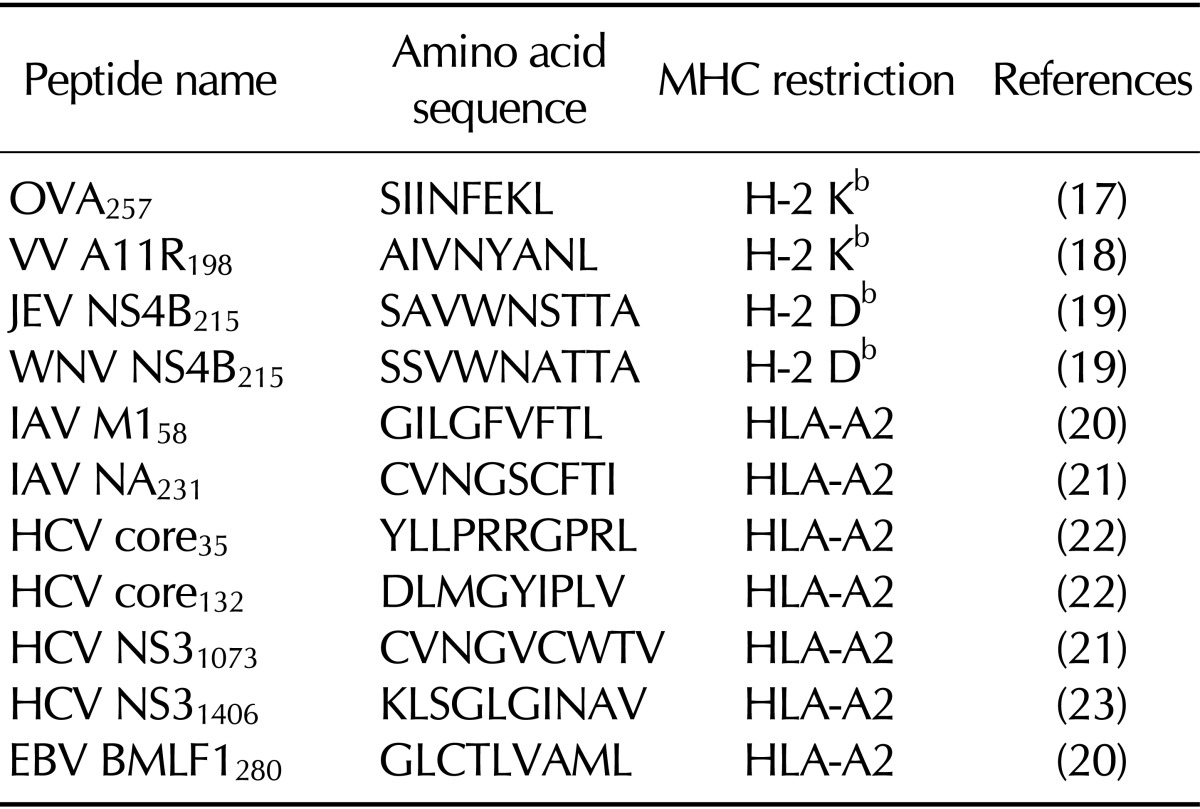
To examine the immunogenicity of H-2 class I-restricted and HLA-A2-restricted peptide epitopes, we immunized HLA-A2-transgenic mice with one of these peptide epitopes and evaluated epitope-specific T-cell responses with direct ex vivo IFN-γ ELISPOT assays. For H-2 class I-restricted peptide epitopes, we used OVA257, VV A11R198, Japanese encephalitis virus NS4B215, and West Nile virus NS4B215 peptides (see Table I). All of these epitopes elicited robust T-cell responses in HLA-A2-transgenic mice (Fig. 1A). For HLA-A2-restricted peptide epitopes, we used IAV M158, IAV NA231, HCV core35, HCV core132, HCV NS31073, HCV NS31406, and Epstein-Barr virus BMLF1280 peptides (Table I). IAV M158, HCV core35, and HCV NS31406 elicited epitope-specific T-cell responses in HLA-A2-transgenic mice, but the other peptides did not (see Fig. 1A). We also performed MHC class I pentamer staining for HLA-A2-restricted peptide epitopes. HCV core132-specific CD8+ T cells were not detected in the spleen after peptide immunization, whereas IAV M158-specific CD8+ T cells were detectable (Fig. 1B), validating the results of the IFN-γ ELISPOT assays. Taken together, these results confirm previous reports that HLA-A2-restricted peptide epitopes tend to be poorly immunogenic in HLA-A2-transgenic mice (13,14,15).
 | Figure 1Immunogenicity of H-2 class I-restricted and HLA-A2-restricted peptide epitopes in HLA-A2-transgenic mice. (A) HLA-A2-transgenic mice were immunized with peptide epitopes, and epitope-specific T-cell responses were evaluated with direct ex vivo interferon gamma (IFN-γ) ELISPOT assays with splenocytes as described in Materials and Methods (n=3-5). (B) HLA-A2-transgenic mice were immunized with peptide epitopes, and MHC class I pentamer staining was performed with splenocytes as described in Materials and Methods. (C) HLA-A2-transgenic mice were infected with recombinant vaccinia virus-hepatitis C virus (rVV-HCV)NS3-4B, and epitope-specific T-cell responses were evaluated with direct ex vivo IFN-γ ELISPOT assays with splenocytes as described in Materials and Methods (n=5; **p<0.01).
|
We also compared the immunogenicity of H-2 class I-restricted and HLA-A2-restricted peptide epitopes in the context of viral infection. HLA-A2-transgenic mice were infected with rVV-HCVNS3-4B, and epitope-specific T-cell responses were assessed with direct ex vivo IFN-γ ELISPOT assays. After rVV-HCVNS3-4B infection, T-cell responses directed to HLA-A2-restricted HCV NS3 peptide epitopes such as HCV NS31073 and HCV NS31406 were scarcely induced, whereas H-2 Kb-restricted VV A11R198-specific T-cell response was vigorously elicited (Fig. 1C). This result shows that HLA-A2-restricted peptide epitopes are not immunodominant in HLA-A2-transgenic mice.
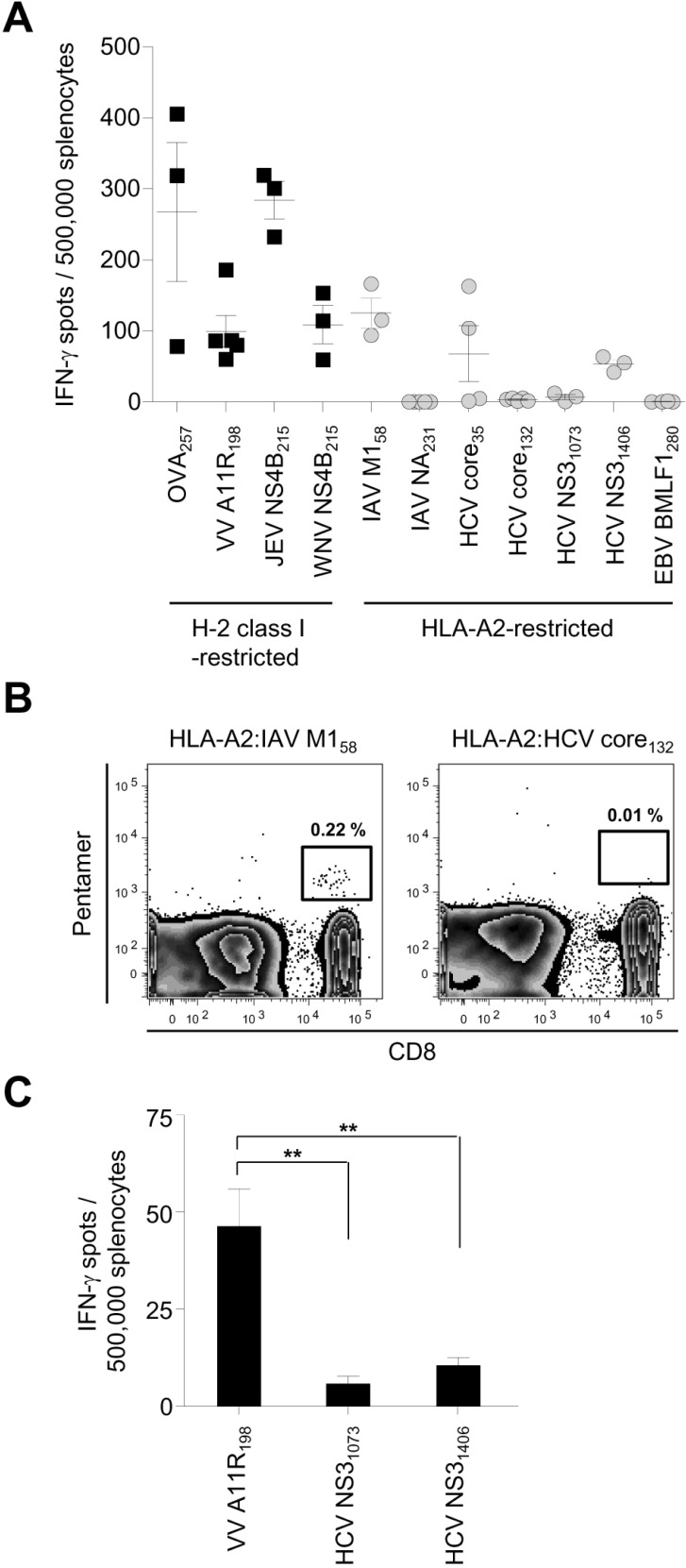
Next, we pursued the explanation for the poor immunogenicity of HLA-A2-restricted peptide epitopes in HLA-A2-transgenic mice by examining the precursor frequency of epitope-specific naïve CD8+ T cells. We isolated cells from the secondary lymphoid organs of the whole body of a mouse, stained them with MHC class I pentamer, and magnetically enriched the MHC class I pentamer(+) cells. After enrichment, CTL precursors recognizing the H-2 Kb-restricted OVA257 epitope were detected in the naïve CD8+ T-cell population of HLA-A2-transgenic mice (Fig. 2A). However, CTL precursors specific to the HLA-A2-restricted HCV core132 epitope were not detected in these mice (see Fig. 2A). In the case of the HLA-A2-restricted IAV M158 epitope that elicited T-cell responses (see Fig. 1A and B), epitope-specific CTL precursors were detected in the naïve CD8+ T-cell population of HLA-A2-transgenic mice (see Fig. 2A). We also calculated the frequency of naïve CTL precursors specific to each peptide in an HLA-A2-transgenic mouse (see Fig. 2A). Taken together, these results led us to conclude that the frequency of naïve CTL precursors is correlated with CTL immunogenicity and the immunodominance of epitopes and that the poor immunogenicity of HLA-A2-restricted peptide epitopes is related to the paucity of naïve CTL precursors in HLA-A2-transgenic mice.
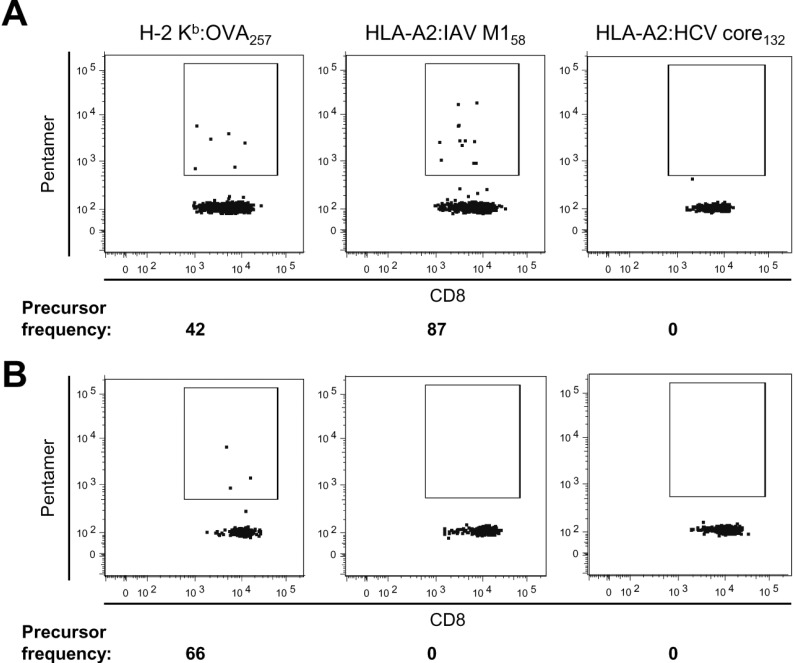 | Figure 2Precursor frequency of epitope-specific naïve CD8+ T cells. Cells were isolated from the secondary lymphoid organs of the whole body of an HLA-A2-transgenic (A) or C57BL/6 (B) mouse and stained with MHC class I pentamer. MHC class I pentamer(+) cells were magnetically enriched and detected with flow cytometry. The precursor frequency of epitope-specific naïve CD8+ T cells was calculated as described in Materials and Methods.
|
We also examined naïve CTL precursors in conventional C57BL/6 mice without the HLA-A2 transgene. As in HLA-A2-transgenic mice, OVA257-specific naïve CTL precursors were detected and HCV core132-specific naïve CTL precursors were not detected in C57BL/6 mice (Fig. 2B). However, IAV M158-specific naïve CTL precursors were not detected in C57BL/6 mice, whereas they were detected in HLA-A2-transgenic mice (see Fig. 2B). These results suggest that IAV M158-specific naïve CTL precursors survive through positive selection in the thymus of HLA-A2-transgenic mice but not in that of C57BL/6 mice. However, the positive selection of HCV core132-specific naïve CTL precursors is not influenced by HLA-A2 expression in the thymus. In this case, the paucity of naïve CTL precursors might be caused by differences in germline TCR genes between humans and mice; there is a hole for TCRs recognizing HCV core132 in the TCR repertoire of C57BL/6-background mice.
Since their establishment, HLA-A2-transgenic mouse strains have been useful tools for the identification of HLA-A2-restricted peptide epitopes within viral or tumor antigen proteins. However, HLA-A2-restricted peptide epitopes tend to be poorly immunogenic in HLA-A2-transgenic mice (13,14,15), although one report has shown an extensive overlap between the TCR repertoires of HLA-A2-transgenic mice and human CTLs (24). In the current study, we demonstrate the poor immunogenicity of several HLA-A2-restricted peptide epitopes in HLA-A2-transgenic mice. To improve the immunogenicity of HLA-A2-restricted peptide epitopes in HLA-A2-transgenic mice, H-2 class I was knocked out in HLA-A2-transgenic mice (13). In fact, HLA-A2-restricted CTL responses in H-2 class I knockout HLA-A2-transgenic mice better resemble human CTL responses (14,15).
Although we examined only HLA-A2-transgenic mice in the present study, we expect similar results in mice expressing other HLA class I allotypes (10,11,12). HLA class II-transgenic mice have also been used for immunological studies (28,29,30,31). Examinations of the immunogenicity of HLA class II-restricted peptide epitopes in relation to the frequency of naïve CD4+ T cell precursors are warranted in HLA class II-transgenic mice, as performed in the current study.
Given the results of the present study, we suggest that differences in germline TCR genes between humans and mice might cause the paucity of naïve CTL precursors recognizing HLA-A2-restricted epitopes in HLA-A2-transgenic mice based on the frequency of HCV core132-specific naïve CD8+ T cells (see Fig. 2A and B). Therefore, there might be holes for TCRs recognizing certain HLA-A2-restricted epitopes in the mouse TCR repertoire. If this is the case, the immunogenicity of HLA-A2-restricted peptide epitopes in HLA-A2-transgenic mice could be improved by the introduction of the human TCR gene loci into HLA-A2-transgenic mice. Then, CTL immunodominance patterns in HLA-A2-transgenic mice would better reflect those in HLA-A2(+) humans. Indeed, mice transgenic for human TCR gene loci have been established, and they have a diverse human TCR repertoire (32). In this way, the T-cell immune system of a mouse model will better resemble that of humans, improving these tools for the study of epitope repertoires and the immunodominance of human T-cell responses.
Go to :
 XML Download
XML Download