

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
IL-33 is a multifaceted, multifunctional cytokine. It was initially found to be highly expressed in the nuclei of endothelial and epithelial cells (1). Binding of IL-33 to a heterodimeric receptor complex consisting of ST2 and IL-1RAP results in the recruitment of an adapter protein, MyD88, and the onset of signal transduction (2). IL-33 signaling induces the production of various mediators involved in inflammation and tissue repair by nonhematopoietic and hematopoietic cells, including endothelial cells, epithelial cells, macrophages, basophils, eosinophils, and mast cells (3).
Candida albicans is a commensal organism of the gastrointestinal tract and vagina (4); however, it is also the most common pathogen associated with mucosal and systemic infections. Invasive candidiasis is the fourth leading cause of nosocomial bloodstream infection in the USA (5), and it is estimated to occur worldwide in over 400,000 people every year with mortalities ranging from 46% to 75% despite administration of antifungal therapy in modern intensive care unit facilities (6). The resistance mechanism of mice to C. albicans infection relies mainly on the phagocytic and killing ability of innate immune cells, such as neutrophils and macrophages (7,8). Our previous studies have shown that IL-33 enhances neutrophil recruitment to site of infection and their phagocytic activities in peritoneal C. albicans infection (9,10). Little is known about the role of IL-33 in macrophages during Candida infection. In this study, we demonstrate that pre-sensitizing peritoneal macrophages with IL-33 enhances various activities that facilitate fungal clearance.
MATERIALS AND METHODS
Mice
C57BL/6 mice were purchased from Orient Bio-Charles River. MyD88 KO mice with a C57BL/6 genetic background were maintained in a specific pathogen-free facility and used when they were 7 to 8 weeks old. All experiments were conducted according to the regulations of the Animal Committee of the University of Ulsan.
Isolation of peritoneal macrophages
Mice were intraperitoneally injected with 3 ml of sterile 3% thioglycollate broth (Difco). After 3 days, mice were sacrificed and peritoneal exudate cells (PECs) were harvested from the peritoneal cavities. The cell pellet was washed and resuspended in DMEM medium supplemented with 10% fetal bovine serum (FBS). PECs were seeded into 24-well tissue culture plate (1×106 cells/ml) and incubated at 37℃ for 2 h, at which point the adherent cells were harvested.
Measurement of cytokines
Cytokines present in culture supernatants were measured by ELISA (eBioscience or R&D systems), according to manufacturers' protocols.
Real-time RT-PCR
Total RNA was extracted from isolated peritoneal macrophages using TRIzol (Invitrogen), according to the manufacturer's manufacturer's instructions. cDNA was synthesized from 2µg of total RNA using SuperScript reverse transcriptase (Invitrogen). Real-time PCR was performed using SYBR Green PCR Master Mix (Qiagen) in the ABI 7500 Fast Real-Time PCR System (Applied Biosystems). The primers used to measure levels of ST2 mRNA were 5'-TGA CGG CCA CCA GAT CAT TCA CAG-3' (forward) and 5'-GCC AAA GCA AGC TGA ACA GGC AAT AC-3' (reverse).
Phagocytosis assay
In vitro phagocytosis assays were performed as previously described (7). Briefly, peritoneal macrophages were incubated with IL-33 (100 ng/ml) at 37℃ for 2 h. Heat-killed C. albicans was labeled with FITC, opsonized, and challenged against IL-33-primed macrophages at 37℃ for 20 min (MOI=10). Phagocytosis was stopped by transferring cells to ice and washing thoroughly with cold FACS buffer. Extracellular fluorescence was quenched by adding 200µl of PBS containing 0.04% trypan blue and 1% formaldehyde. Fungus-containing cells were then counted by flow cytometry and the degree of phagocytosis expressed as the percentage of FITC-positive macrophages.
Killing assay
Macrophages were mixed with opsonized live C. albicans (MOI=1) and incubated at 37℃ for 20 min with continuous rotation. Cells were washed thoroughly in cold PBS, resuspended in warm DMEM medium, and further incubated at 37℃. At the indicated times, 200 ml samples were harvested and cells lysed in PBS containing 0.1% Triton X-100. CFUs were quantified by plating lysates on agar. Percent killing was calculated as [1-(CFUs after incubation/phagocytosed CFUs at the start of incubation)]×100.
Determination of reactive oxygen species (ROS) generation
Macrophages were pre-sensitized with IL-33 (100 ng/ml) or PBS for 2 h and were then further incubated in the presence of 2µM 2',7'-dichlorodihydrofluorescein diacetate (Molecular Probes) for 20 min in the dark. Cells were washed twice with PBS, challenged with heat-killed C. albicans (MOI=10) for 10 or 30 min, and analyzed by FACS.
Flow cytometry
Prepared cells were blocked with 2.4G2 mAb in staining buffer (PBS containing 0.2% BSA and 0.1% sodium azide) at 4℃ for 20 min, incubated with rat anti-mouse T1/ST2 (clone DJ8)-FITC mAb or rat IgG1κ isotype control at 4℃ for 20 min, and then washed twice with staining buffer. Flow cytometric analysis was performed using a FACS Canto II unit (BD Biosciences). Data were analyzed with FACS Diva (BD Biosciences) and FlowJo software (TreeStar).
RESULTS
IL-33 enhances the production of pro-inflammatory cytokines by macrophages via the ST2/MyD88 signaling axis
To investigate whether IL-33 can affect cytokine production by macrophages in response to C. albicans, peritoneal macrophages were stimulated with heat-killed Candida (MOI=10). We found that C. albicans markedly increased the secretion of IL-6 and TNFα by macrophages at 6 h after challenge and thereafter (Fig. 1). Pre-sensitization of macrophages with IL-33 enhanced the production of these pro-inflammatory cytokines following challenge with C. albicans (Fig. 1).
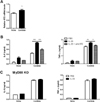
Since IL-33 signaling is mediated by ST2 (11), we examined whether the expression of ST2 in peritoneal macrophages is regulated by C. albicans. RT-PCR analysis demonstrated that ST2 expression was significantly increased in macrophages 24 h after stimulation with C. albicans (Fig. 2A). We next sought to determine whether the pre-sensitization effect of IL-33 was dependent on ST2. Notably, pre-treatment with a ST2 neutralizing antibody abolished the effect of pre-sensitization with IL-33 on IL-6 and TNFα production following C. albicans infection (Fig. 2B). As ST2 signaling is dependent upon MyD88, we further analyzed the effect of pre-sensitization with IL-33 on pro-inflammatory cytokine production in MyD88 KO peritoneal macrophages. As expected, IL-33 pre-sensitization had no effect on the secreted levels of IL-6 and TNFα following C. albicans challenge in MyD88 KO macrophages (Fig. 2C). These data clearly establish that the effect of pre-sensitization with IL-33 on IL-6 and TNFα production by peritoneal macrophages after C. albicans challenge is mediated through the ST2-MyD88 signaling axis.
IL-33 pre-sensitization enhances the phagocytic activity of macrophages
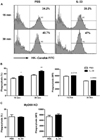
We have previously shown that IL-33 pre-sensitization can increase the phagocytic activity of neutrophils against C. albicans (9,10), though little is known about the effect of IL-33 on macrophage phagocytosis. As seen in Fig. 3A, IL-33 priming results in a significant increase in the overall percentage of macrophages containing FITC-conjugated C. albicans at 15 and 30 min post-challenge, yet this was accompanied by only a marginal increase in the FITC intensity observed in C. albicans-engulfed macrophages (Fig. 3B). Notably, IL-33 priming had no effect on the phagocytic activity of MyD88 KO peritoneal macrophages against C. albicans (Fig. 3C). These results indicate that IL-33 priming enhanced macrophage phagocytosis in a MyD88-dependent manner by expanding the population of phagocytic macrophages targeting C. albicans, rather than elevating the phagocytic ability of the individual macrophages.
In addition to phagocytosis, the ability of macrophages to kill engulfed pathogens is also critical for the innate immune response to fungal infection (12,13). Notably, the capacity to generate and utilize intracellular reactive oxygen species (ROS) is crucial for killing C. albicans in phagocytes (14,15). To determine the role of IL-33 priming on ROS production, we analyzed the fluctuations of ROS levels in IL-33-primed peritoneal macrophages in response to fungal challenge. As shown in Fig. 4A, IL-33-primed macrophages displayed higher intracellular ROS levels following challenge when compared to mock-primed controls. Consistent with this result, phagocytosed C. albicans were more rapidly cleared inside IL-33 primed macrophages (Fig. 4B).
DISCUSSION
We have previously shown that administration of IL-33 prior to peritoneal challenge with a lethal dose of C. albicans prevents sepsis-induced mortality (9,10). In that model, pre-sensitization of the host with IL-33 increases neutrophil responses at multiple stages. Notably, IL-33-primed peritoneal macrophages play a critical role in neutrophil recruitment by producing the CXCR1/2 necessary for neutrophil chemotaxis. The data presented in this study further indicate that peritoneal macrophages may be critical in neutrophil activation, as IL-33-primed macrophages are sufficient to provide the pro-inflammatory cytokines prerequisite for full neutrophil activation. Our results also suggest that IL-33-primed macrophages may contribute to fungal clearance by directly phagocytosing C. albicans, though in vivo experiments are necessary to confirm our in vitro observations.
Although neutrophils play a key role in the anti-fungal defense against C. albicans, macrophages appear to be of equal importance in fungal clearance (16,17). A recent study demonstrates that treatment with IL-13 or PPARγ ligand enhances dectin-1 receptor expression, resulting in induction of phagocytic activity in peritoneal macrophages (18). Interestingly, IL-33 can strengthen M2 macrophage activation to enhance fungal clearance in response to Pneumocystis murina (19). Similarly, our unpublished data revealed that IL-33 pre-treatment can result in a significant increased M2 macrophage polarization in the kidney following C. albicans systemic infection at 3 d post-challenge and thereafter; however, the majority of macrophages present in the kidney at 1 d post-infection display a M1 phenotype. Taken together, this suggests that IL-33 promotes the activation of M1 macrophages during early C. albicans infection, and then slowly converts them toward a M2 macrophage phenotype. Nevertheless, both types of macrophages are known to mediate activities necessary for fungal clearance. This characteristic of IL-33 is unique among cytokines and has a merit in its ability to enhance both resistance and tolerance to C. albicans infection (our unpublished data). In summation, our in vitro data indicate that IL-33 may function as an important mediator of anti-fungal host defenses early after C. albicans infection by promoting fungal clearance.
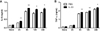

 XML Download
XML Download