

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
CCL
chemokine (C-C motif) ligand
CDM
limonoid 1-cinnamoyl-3,11-dihydromeliacarpin
CNS
central nervous system
CTL
cytotoxic T lymphocyte
CXCL
chemokine (C-X-C motif) ligand
DCs
dendritic cells
gB
glycoprotein B
HSV-1
herpes simplex virus type 1
HSV-2
herpes simplex virus type 2
HIV
human immunodeficiency virus
LCs
langerhans cells
LPS
lipopolysaccharide
MAPKs
mitogen-activated protein kinases
M-CSF
macrophage-colony stimulating factor
MIP-1α
macrophage inflammatory protein-1 alpha (CCL3)
MyD88
myeloid differentiation primary response gene 88
Poly-IC
polyinocinic-polycytidylic acid
PRRs
pattern recognition receptors
PAMPs
pathogen associated molecular patterns
TCR
T cell receptor
TLRs
Toll-like receptors
INTRODUCTION
Herpes simplex viruses (HSV), the most prevalent and pestilent causes of human viral infections, belong to the genus simplexvirus of the family Herpesviridae including two members herpes simplex virus 1 and 2 (HSV-1 and HSV-2). Genome of HSV is linear double-stranded DNA being enclosed with enveloped icosahedral capsid. The envelope holds at least 10 different glycoproteins protruding from the outer side (gB, gC, gD, gE, gG, gH, gI, gK, gL, and gM), which have their primary mechanical functions in viral attachment and entry as well several immune regulatory effects. The immune response against HSV involves intricate and multifactorial; yet these viruses have various immune evasion and modulation mechanisms that are resulted in their evolutionary success. On that ground, the study of anti-HSV immune responses, including both innate and adaptive immune responses as well as the corresponding viral oppugn measures, is valuable to our understanding of HSV pathogenesis and anti-HSV immunity (1,2,3,4).
The innate antiviral response is account to play a vital role in determining the outcome of an HSV infection. The first line of defense against HSV1/2 infection is provided by innate humoral (complement, cytokines, chemokines) and cellular (interferon-producing cells, macrophages, neutrophils, NK cells and γδ T cell) responses. These orchestrate the lysis of virions and virus-infected cells and subsequently provide a link to effective adaptive immunity, too (3,4,5,6). The adaptive immune system, also called the acquired immune system, has been shown to play important roles in disease progression, latency and control of virus spread. Key components of the adaptive immune system are B-cells, T-cells, antibodies and the secondary lymphoid organs (7,8,9,10).
Developing novel strategies to eradicate HSV is a global public health priority. There are three ways to eradicate HSV infections in terms of anti-HSV drugs, microbicides and vaccines (Summarized in Table I). Prevention is better than cure as antiviral drugs do not destroy their target pathogen; instead they inhibit their development. There are three classes of drugs currently licensed for the treatment of HSV infection, all of which target viral DNA replication; these include guanosine analogues (acyclovir, famcilovir, and ganciclovir), acyclic nucleotide analogue, cidofovir and the pyrophosphate analogue, foscarnet. A microbicide is any biocidal compound or substance whose purpose is to reduce the infectivity of microbes, such as viruses or bacteria. A novel strategy for HSV prevention is the development of mucosally delivered microbicides. Historically, prevention acquired by vaccines is considered as the best way to control the HSV epidemic. The development of vaccines/biologicals that elicit sterilizing immunity to completely prevent the establishment of infection ultimately results in prevention and eradication of infection (11,12,13).
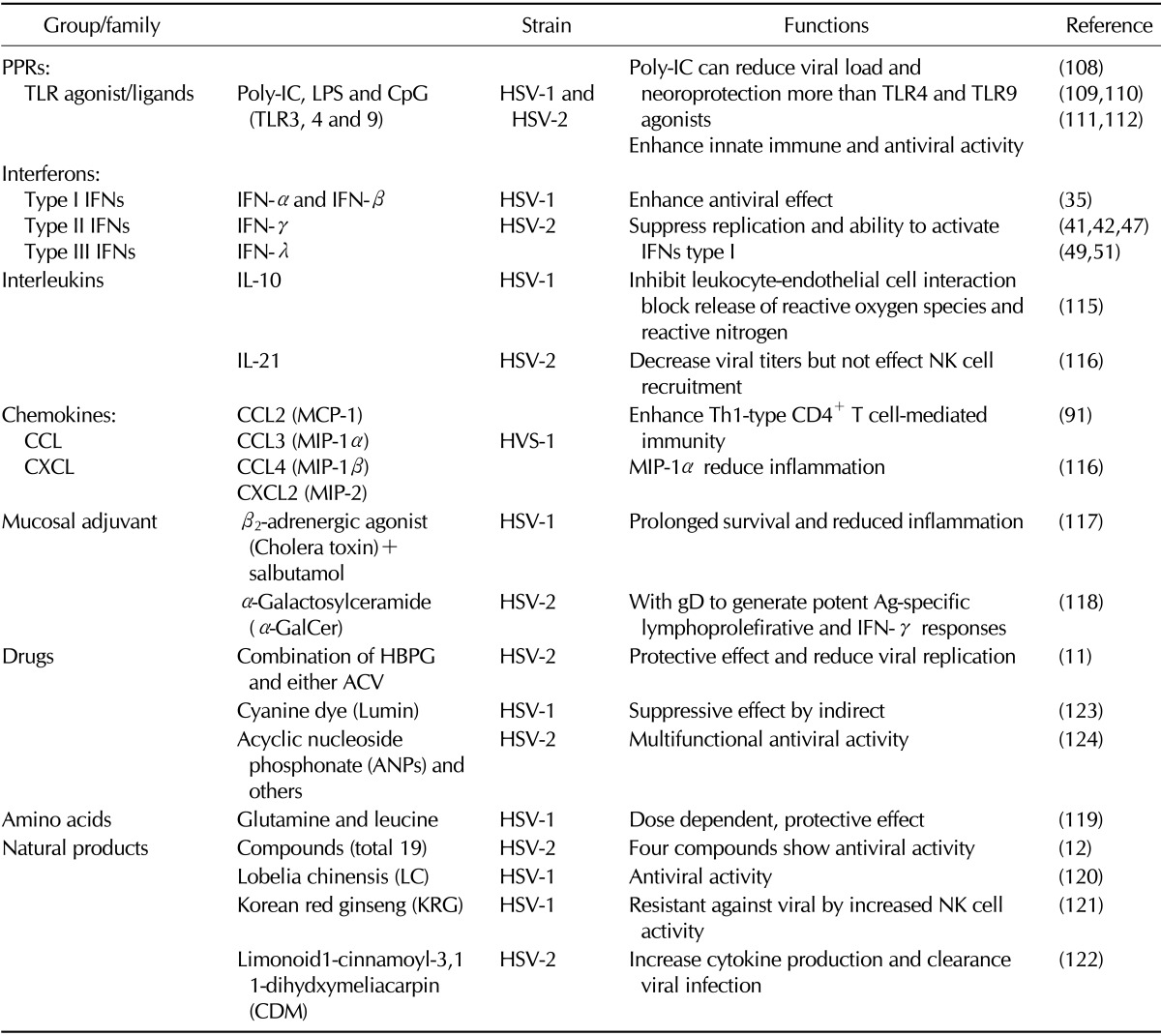
This review provides an update on researchers regarding the modulation on innate and adaptive immunity against HSV, which is focused on mucosal route of infection. In addition, current status of treatment and prophylactic measures for development against both HSV-1 and HSV-2 was discussed. Also, here focus on how immune system modulation help in developed novel strategies regarding drugs, microbicides and biologicals to eradicate HSV.
Go to : 
MODULATION OF INNATE IMMUNITY AGAINST MUCOSAL INFECTION OF HSV
The innate or non-specific immune system is one of the major subdivisions of immune system. The innate immune system is our first line of defense against invading organisms. The elements of the innate (non-specific) immune system include anatomical barriers, secretory molecules and cellular components.
Importance of TLR signaling against HSV mucosal infection
Toll-like receptors (TLRs) are important element from innate immunity. TLRs are membrane-bound pattern recognition receptor (PPR) proteins expressed by cells of the innate immune system to proclaim pathogen associated molecular patterns (PAMPs). TLRs are single transmembrane non-catalytic receptor protein which plays informational role in innate immunity network against microbial pathogens and successive induction of adaptive immunity. All TLRs possess amino-terminal leucine-rich repeats, which are responsible for the recognition of PAMPs, and also possess a carboxy-terminal Toll-interleukin-1 (IL-1) receptor (TIR) domain, which is required for initiating intracellular signaling (14,15). TLRs interact with their specific PAMP to induce NF-kB signaling and the MAP kinase pathway, and subsequently led to the secretion of proinflammatory cytokines and co-stimulatory molecules. Although engagement of each TLR activates a different specific molecular cascade, many induce the production of Th1-type cytokines, such as IL-12 and IFN-γ (16). These molecules act as activation signal to other cells of the immune system making TLRs as key link between innate immunity and adaptive immunity (17).
TLRs had been first discovered in Drosophila and subsequently have been found in many species. There are 10 human (TLRs 1 to 10) and 12 murine (TLR1 to 9 and TLR11 to 13) TLR family members. TLR3 and 7 to 9 recognize PAMPs in endosomes and all remaining TLRs in extracellular space (14). There are several TLRs monitors, especially TLR2, TLR3, and TLR9, involved in early recognition of HSV components (18). TLR2 recognizes an unidentified molecular structure on the virion. A number of studies revealed that TLR2 mediates the induction of inflammatory cytokines response to HSV-1, but that expression of TLR2 is not protective against lethal viral encephalitis on HSV-1 infection (19,20). In vitro study demonstrates the importance of TLR2 in microglial cell to induce oxidative stress through decreasing activation of p38 MAPK and p42/p44 ERK and neuronal damage in response to HSV-1 infection (21). TLR9 recognizes unmethylated CpG sequences in DNA molecules. One study result indicates that TLR9 mediates cellular response to unmethylated CpG dinucleotides in bacterial DNA and viral DNA to mount an innate immune response (22). Some studies showed that TLR9 plays an important role for dendritic cell (DC) response to HSV-1, but they did not find differences in viral replication or in susceptibility in TLR9- and MyD88-deficient mice infected with HSV-1 via the footpad or the cornea route (23,24,25). Herein, some studies have shown the secretion of type I IFN in response to HSV in vivo as well as in vitro mediated by TLR9/MyD88-dependent and independent pathway (23,25). However, others demonstrated that MyD88-deficient mice showed fully 100% lethal encephalitis when infected via mucosal route (i.e. intranasal and intravaginal) with HSV-1 and HSV-2, respectively (2,26).
TLR3 is efficient in the recognition of double stranded RNA set up during the virus replication. About TLR3 signaling, some study suggested that TLR3 provides early control of HSV-1 and 2 infections immediately after entry into the CNS by mediating type I IFN responses in central nervous system (CNS) (18,27,28).
The role of type I, II and III IFNs in mucosal infection of HSV
The interferon system is one of the major elements from the innate immune system, which is the first line of defense against various viral infections in mammals. This system is designed to block the spread of virus infection in the body, sometimes at the expense of accelerating the death of the infected cells. Interferons (INFs) are cytokines that cause cells to limit or prohibit viral replication, and they have a variety of functions in the innate immune response. They can control macrophage and NK cell activation, stimulate cytotoxic lymphocytes, induce cell surface costimulatory molecules, activate cytokine production, and stimulate local inflammation (3,4,29). IFNs were described and named in 1957 by Alick Isaacs and Jean Lindenmann (30). Seven IFNs have been described for humans out of ten distinct IFNs identified in mammals. They are typically divided among three IFN classes: type I IFN, type II IFN and type III IFN. All IFN classes are very important for fighting viral infections and tumors (31). The type I IFNs in humans are IFN-α, IFN-β and IFN-ω and bind to common receptor known as IFN-α receptor (IFNAR) that consists of IFNAR1 and IFNAR2 chains. In human type II IFNs is IFN-γ and signals intracellular transduction through IFN-γ receptor (IFNGR) complex that consists of IFNGR1 and IFNGR2 chains (32). Types I and III IFNs were produced by most cell types in particularly pDCs and cDCs (30,31,33,34) and INF-γ is produced by natural killer (NK) cells and CD4+ T cells, CD8+ T cells (3,4,6,7).
Many various studies are focused on the role of type I and II IFNs during the HSV-1 and HSV-2 infection, respectively. Initially, Gill et al. and others demonstrated that TLR-mediated protection is dependent on type I signaling (35,36). Importantly, recent data suggested that IRF-3 activation and subsequent IFN-α/β signaling are required for poly (I:C)-induced innate protection against intravaginal HSV-2 challenge. Collectively, local delivery of murine recombinant IFN-β alone was shown to protect C57BL/6 and IRF-3-/- mice against subsequent intravaginal HSV-2 challenge (35). Also, Conrady et al. clearly showed the importance of IFN production in innate immune response as well as evoking several chemokine's production necessitated to assist adaptive immune response in response to both HSV-1 and HSV-2 infection via ocular and intravaginal route (37,38,39). More recently, one study demonstrated that type I IFN induced the production of IL-15, which promotes NK cell survival and proliferation during the HSV-2 infection (40).
The antiviral effect of IFN-γ may be direct (intracellular, NO secretion) or indirect, involving activation of effector cells of the immune system. Several studies were focused on the type II IFN (IFN-γ). Initial some studies reported that IFN-γ is important for controlling in vivo-reactivated HSV-1 and thereby contributes to the maintenance of virological latency, meaning the absence of infectious HSV-1 in the ganglion as opposed to molecular latency, which is manifested as repression of viral gene expression at the cellular level (41,42). Additionally, one study found that both IFN-γ and T cell-mediated cytolytic mechanisms which are mediated by either perforin or Fas are required for complete clearance of HSV-2 from the genital epithelium (43). However, contrast one research team suggested that IFN-γ is not fully helpful for virus clearance but plays a key role in enhancing T cell immune response in HSV-1 reactivation (44).
Type III IFN, also known as interferon lambda (IFN-λ), is recently classified subfamily of interferon. IFN-λ1-3 or IL-28A/B and IL-29 are three structurally related members of this group. They signal through a receptor complex consisting CRF2-4 (IL-10Rβ) and CRF2-12 (IFNLR1) (33,34). A number of studies suggested the role of type III IFN during HSV infection. It has been shown to have antiviral activity in vitro as types III IFNs trigger type I IFN-like gene expression profile (45,46,47). Ank et al. shows that IFN-λ produced by DCs at mucosal sites elicited limited antiviral activity to HSV-2 infection with its stronger dependence on NF-κB through TLR9, compared to IFN-α (48,49). After that, Marie et al. reported that DCs are a key source of IFN-λ at epithelial surfaces in the vaginal tract and that expression of type III IFN show high level of dependent on the NF-kB pathway than type I IFNs (50). Also, Jieliang et al. showed that the replication of HSV-1 infection is suppressed in microglia and astrocytes by treatment of IFN-λ through TLR3 activation (51,52).
NK cells, macrophages, and dendritic cells
NK cells, monocytes, neutrophils, macrophages and DCs are innate immune cells. They play a crucial role during the early phase of a viral infection. NK cells are important cellular component of the innate immune response against tumor cells and virus-infected cells and to produce cytokines such as IFN-γ and granzyme B (3,4,5,53). Studies clearly demonstrated that cytokines, such as IL-2, IL-12, IL-15, and IL-18, involved in NK cell activation (40,54,55,56). For instance, Ashkar and Rosenthal clearly demonstrated that NK, NKT and IL-15-ablated mice were very high susceptible to low dose (100 PFU) challenge and showed 100% mortality, compared to B6 control mice during HSV-2 infection via vaginal route (55). Also, IL-18 plays a key role in the rapid activation of NK cells, thereby resulting in control of early HSV-1 replication in the lung in intranasal HSV-1 infection (56). Also others result suggested that type I IFNs signaling are key mediator for NK cell activation by IL-15-help during genital HSV-2 infection (40). Several studies have suggested that NK cell depletion via anti-NK1.1 or anti-asialo-GM1 antibodies increases the susceptibility of mice to ocular, genital, cutaneous and intravascular challenge with HSV (40,53,57,58). Importantly, recent one study also revealed that NK cells are stimulated by HSV Ag through TLR2 activation, and subsequently can contact with CD4+T cells in direct cell-to-cell manner (6).
DC is professional antigen-presenting cells and bridges between the innate and adaptive immune system (59). Although many studies focused on DC against HSV-1 and HSV-2, initially Zao et al. demonstrated that vaginal submucosal CD11b+ DCs, but not Langerhans cells (LCs), induce protective role of Th1 responses in the draining lymph nodes (60). Next others using mucosal viral infection, it is shown that viral recognition and induction of antiviral immunity by both the infected stromal cells and uninfected Ag-presenting DCs requires TLR signaling (61). HSV-1 entry glycoproteins act as a target for innate immune recognition that is going to activate DCs independenting on TLR2 signaling (62). Sadik et al. suggested that DCs are essential not only in the optimal activation of NK cells and CD4+ and CD8+ T cells but also need for resistance to HSV-1 infection (63). Following early HSV-1 corneal infection, resident DCs, but not polymorphnuclear neutrophils, had essential role for migration of NK cells and inflammatory monocytes into the central cornea (64). Interestingly herein, one research team suggested that PD-1: PD-L1 signaling produced by DC mediates T-cell exhaustion and latency during acute ocular HSV-1 infection (65). Additionally, several research teams tried to clearly describe the role of plasmacytoid DCs (pDC) during HSV mucosal infection. pDCs have been identified as a potent secretor of the type I IFNs in response to CpG as well as several viruses. Therefore, Lund et al. and other research teams demonstrated that pDC can recognize both HSV-1 and HSV-2 exclusively via TLR9 (66,67,68).
Macrophages are anti-herpetic actions during the first hours of the infection. Besides that, macrophages also play wide variety of immune functions which are including phagocytosis, tumor cytotoxicity and secretion of cytokine and antigen presentation (3). Mott et al. clearly suggested that STAT1 may have critically important role for allowing bone marrow-derived DCs and macrophages to blocking HSV-1 replication (69). Present study hypothesized that macrophage treated with IL-27 might induce the expression of novel micro RNAs that may be affecting the anti HSV function in IL-27 and M-CSF (I-Mac) (70). Also, recent study recognized that ablation of macrophages, but not DCs, NK cells, B cells, CD4+ T cells, or CD8+ T cells, induced CNS demyelination during the infected ocular with HSV-1. Finally, their results showed that macrophage IL-12p70 signaling could inhibit development autoaggressive CD4+ Treg cells, resulting in the prevention of HSV-1 induced CNS injury (71). More importantly, others demonstrate that macrophage-mediated immunity against infection occurs efficiently through iNOS in trigeminal ganglia and appears to be organized by TLR2 and TLR9, which contributes to HSV-1 infection control (72). Also, inflammatory monocytes were found to play main role in antiviral defense against HSV-2 genital mucosa infection through CCL2 molecule (73). Neutrophils also appear to have a key role in T and Bcell recruitment and control of viral replication during both HSV-1 and HSV-2 in corneal infection and vaginal mucosa (74,75). But constantly, recent one study suggested, using depletion model of Gr-1+ cells, that neutrophils may be do not play a major role in HSV-1 clearance in an intranasal model (76).
Go to :
MODULATION OF ADAPTIVE IMMUNITY AGAINST MUCOSAL INFECTION OF HSV
The adaptive immune response against pathogen is primarily responsible for viral clearance and the generation of long-term memory. Current vaccine development strategies have focuses on understanding adaptive cellular-mediated immune responses. The effective adaptive responses predominantly require a balance between CD4+ and CD8+ T-cell activation. Acquired protective immunity can be mediated by both humoral and cell-mediated immune mechanisms.
Is humoral immunity indispensable for the clearance of HSV?
Many studies showed that B cells producing natural antibodies including IgA and IgG have been shown to have anti-viral and inflammatory effects and can control both HSV-1 and HSV-2 infection via vaginal route (7,77,78,79). Additionally, several research teams tried to determine the role of B cells using human and mouse model in ocular infection (80,81). Iijima et al. suggested that B cells together with DC contribute to restimulate memory CD4+T cells to secrete IFN-γ (82). In general, vaccine against HSV was focused on the production of neutralizing antibodies against HSV several glycoproteins (83,84,85). On the other hand, recent one researcher team reported that HSV-1 induced humoral response, especially induction of memory B cells, is dependent on complement system (9).
Although many studies have demonstrated the protective role of B cells against HSV infection, there is also contrasting reports that B cells are not required for HSV-1 and HSV-2 clearance (86,87,88). Therefore, it is still not clear about the specific contribution of humoral immunity to HSV control. While antibodies against HSV can mediate prophylatic protection in mice, B cells are not absolutely required for protection in the context of an acute infection; rather, they likely interact with other immune effectors such as T cells. Taken together, a better understanding of this area of HSV immunity is warranted.
T cell-mediated immunity to HSV mucosal infection
Prominent role of Th1-type CD4+ T cells to HSV mucosal infection
IFN-γ is signature cytokine produced by CD4+Th1 cells which are derived by IL-12 through T-bet transcription factor, and promote cell-mediated immunity against intracellular pathogens (89,90). Several studies showed that IFN-γ-secreting CD4+ T cells are crucial for development of protective immunity against to genital and ocular HSV-1 and HSV-2 infection, respectively (6,7,41,91). Also, Kuklin et al. clearly suggested that CD4+ T cells act as the principal mediators of vaginal immunity against HSV-1 (86). The local NK cells are likely to contribute to early control of HSV replication in infection and make some IFN-γ in the infected tissue (6). Importantly, Kumamoto et al. and other revealed that CD4+ T cells support and help cytotoxic T lymphocyte priming and generation (92).
Some research teams were interested and tried to determine the relative impact of both CD4+ T cells and CD8+ T cells during HSV infection. Herein, against ocular HVS-1, one study showed that both CD4-deficient and CD8+ deficient mice significantly developed corneal scarring compared than control C57BL/6 mice. Also, virus clearance from the eyes of the CD4-deficient mice was longer than CD8-deficient mice (93,94). Additionally, Koelle et al. suggested the clearance of HSV-2 from recurrent genital lesions correlates with the infiltration of both HSV-2-specific CD4+ and CD8+ cytotoxic T cells (95). More importantly, memory Th1 cells, but not CD8 T cells, are localized in the vaginal mucosa and are required for virus clearance after HSV-2 secondary challenge (41).
Paradoxical role of CD4+Foxp3+ Treg and IL-17+CD4+ Th17 cells to HSV mucosal infection
For determining the role of Treg in the control of HSV infections some publications focused on the suppressive role of Treg on CD4+ and CD8+ T cells (96,97). Notably, herein one study showed that adoptive transfer of in vitro-converted CD4+CD25+Foxp3+ Treg cells could decreased lesion severity in initial phase of three different models of herpetic stromal keratitis (HSK) (98). On the other hand, present study found in the female genital tract of mice that TCRγδ+CD4-CD8- T cells are the major population of IL-17A-secreting cells and TCRγδ+ T cells exhibit different expression profiles of cytokines and transcription factors compared to those from spleen (99).
Cytotoxic CD8+ T cells for viral clearance
Rapid induction of CD8+ cytotoxic T lymphocyte (CTL) responses is critical to combat acute infection with intracellular pathogens. Initial study suggested that cross presentation is main roles for induction of CTL to the Ag presentation capacity of CD8α+ DC (100). Also, one study tried to define about HSV epitopes that are recognized by CD8+ T cells in BL/6 mice, and found that nearly 50% of CD8+ T cells participate in recognizing HSV-1epitope (101). In summary, several studies data suggested that CD8+ T cells are require for complete clearance of HSV-1 and 2 (42,43,102,103), whereas CD8+ T cells are dispensable to control virus replication in the vaginal mucosa and other infected area (82,86). Additionally, in latent infection, augmenting the number of circulating HSV-specific CD8+ T cells is not sufficient to bolster the HSV-specific memory T cell population in sensory ganglia (104).
Go to :
PROPHYLACTIC AND THERAPEUTIC STRATEGIES TO HSV MUCOSAL INFECTION
The development of effective prophylactic and therapeutic against herpes mucosal infection still has proven problematic. Difficulties are associated with the complexity of the virus life cycle (latency) and our relatively poor understanding of the mechanism of immune control of primary and recurrent disease (105). The morbidity and socioeconomic burden associated with genital herpes as well as the alarming relationship between genital herpes and the increased risk of acquiring a HIV infection emphasize the need for development of an effective vaccine and/or therapeutics. Primary HSV exposures first elicit innate immune responses and therefore, immunomodulatory approaches are being explored that boost innate immunity and engender increased resistance to HSV infection (91,106,107). Regarding the TLR expression profile in the female genital tract, immunoprophylactics and therapies targeting on TLR2, 3, 7 and 9 of the mucosa have been investigated for the utility to prevent or attenuate herpes virus disease. For prophylaxis, these synthetic agonists were designed to transiently activate the innate immune response to establish a more HSV-resistant environment, thereby increasing the threshold of infection or attenuating recurrent shedding events (108). Recent some works have also suggested that TLR3 agonist (poly: IC) induces more potent antiviral response than the agonists of TLR4 and TLR9 in genital HSV-2 and HSV-1 encephalitis, respectively (109,110). Additionally, some studies tried to determine role of CpG during mucosal HSV infection (111,112). For instance, one study has shown that CpG of HSV genome directly activates pDCs to induce the expression of IFN-α (113). Interestingly, herein one study suggested the mucosa delivery of CpG oligodeoxynucleotides expands functional DCs and macrophages in the vagina (114).
Cytokines may exert antiviral effects via either directly their receptors or indirectly immune modulatory effects. The efficacy of cytokine therapies have been demonstrated in several human and animal studies against HSV infection. It has been already suggested that IFN treatment could be effective to inhibit virus infection and spread in HSV-1 (42,51). Also, others results tried to determine role of some chemokines during HSV infection. Tumpey et al. indicate that IL-10-mediated suppression of MIP-1α synthesis was a significant factor reducing inflammation in the HSV-1 infected cornea (115).
Alteration of innate immunity to viral infectious using treatment with mIL-21 resulted in decreased vaginal viral titers, but did not affect NK cell recruitment and did not subsequently alter IFN-γ production or degranulation activity in vaginal NK cells in the absence of the IL- 21R after intra-vaginal HSV-2 infection (116). Others also examined effects of murine chemokine DNA which are including CC chemokines macrophage inflammatory protein 1β (MIP-1β) and monocyte chemotactic protein 1 (MCP-1), as genetic adjuvants given mucosally. Their results indicated that chemokines function may be displayed by affecting the interaction between innate and adaptive immunity during the HSV-1 infection (91). Moreover, co-administration of salbutamol with DNA vaccine could provide the effective and rapid responses to HSV-1 mucosal challenge, thereby conferring prolonged survival and reduced inflammation against viral infection (117). Another one team also tried to determine effect of α-GalCer, which is mucosal adjuvant and they found protective immunity by intravaginal immunization against vaginal HSV-2 challenge (118). Next, administration of amino acids such as Glutamine and Leucine, but not their combination, showed enhanced production of IFN-γ by NK and suggested that Th1 type CD4+ T cells may be critical to control the outcome of disease following HSV-1 mucosal infection (119).
Since natural products are considered powerful sources of novel drug discovery and development against pathogen. Therefore, many studies investigated the protective effect and immune modulatory of those natural products such as plant extracts against mucosal herpes infection (120,121). Importantly, initial results demonstrated that four compounds, especially eugenol, carrageenan lambda type IV, cineole and curcumin, provide the significant protection than others totally 19 such compounds both in-vitro and in-vivo (12). Interestingly, just few months ago, one research team tried to determine effect of one natural product which is isolated from leaf extract named by limonoid 1 - cinnamoyl - 3, 11 - dihydroxymeliacarpin (CDM) which can inhibit HSV-2 multiplication in epithelial cells and also increases cytokine production in macrophages, both important actions to the clearance of infecting virus in the mouse vagina (122).
There are a variety of nucleoside analog drugs used to treat herpes infections, though the person still harbors the virus for life. Some of these drugs are very specific and are only activated by specific viral enzymes, meaning that these drugs show few side effects such as acyclovir, famciclovir and valacyclovir, and these drugs act against the replicating virus and therefore they are ineffective against latent virus (11,12,122). One report suggested that thymidine kinase inhibitor (2-phenylamino-6-oxo-9-(4-hydroxybutyl)purine; HBPG) may have synergistic activity against HSV encephalitis (11), but others results showed, that cyanine dye (lumen) exhibits significantly suppressive effects on human amnionic FL cell line by suggesting the inhibition of virus invasion into the cell. Thus, that may potential new preventive anti-herpetic drug together with IFN (123).
The relationship between genital herpes and the increased risk of acquiring a HIV infection emphasize the need for development of an effective vaccine and/or therapeutics. Especially, important task is to develop a compound that is highly potent against both viruses to suppress their transmission and replication (91). Herein, Jan Balzarini et al. was focused on a distinct new subclass of acyclic nucleoside phosphonate (ANP), which are structurally and functionally different from previously used drugs such as tenofovir and adefovir that has significant relevance over the commonly used drugs. In their study they use representative drug PMEO-DAPym decisively suppresses HSV DNA polymerase and at the same time the drug also activate anti HIV CC chemokine. This combine results into dual anti-viral therapy (124).
Existing control measures against various infectious diseases includes the combined use of vaccines, antibiotics and chemicals. Vaccines can provide long term immunity and thereby confer specific protection against a particular pathogen following immunization. Studies on efficacy of various vaccines that prevent or reduce the primary and recurrent HSV infection have demonstrated the importance of cellular immunity for protection against the infection. Generally, against HSV vaccine strategies within the last few years have focused on the use of HSV viral epitopes such as gB, gC, gD and gG (77,78,83,84,85,125,126), some specific peptides (127,128), DNA-based vaccine or plasmid viral vectors (129) and attenuated/replication-detective versions of both HSV-1 and HSV-2 respectively (130,131). For example, in human, glycoprotein D-based subunit gD-2 vaccines can provide effective protections in HSV-1 but not HSV-2 seronegative women (132). Additionally, several studies suggested that subunit vaccines in combination with adjuvant appeared to be safe and effective against genital herpes in guinea pigs (133,134), but in clinical trials still failed to provide general protection (135). Importantly, present study demonstrated that vaccination with HSV-1 recombinant CJ9-gD elicits strong and protective immune responses against primary and recurrent HSV-2 genital disease and significantly reduces the extent of latent infection (136). Also, another team tested gB1 vaccine delivered by feline immunodeficiency virus (FIV) vector to elicit cross-neutralizing antibodies and cell-mediated responses that protected 100% and 75% animals from both HSV-1 and HSV-2 associated severe disease, respectively (137). Interestingly, herein one research team tried to determine role of DC for vaccine design and employed to assess their value in protection against live virus challenge in an experimental model using lethal and latent herpes simplex virus (HSV) infection in Balb/c mice (138).
Go to :
CONCLUSIONS AND PERSPECTIVES
Generally, the immune response involves multiple mechanisms to effectively clear viral infection. The innate immune response is the host's first line of antiviral defense, and is mediated through the production of type I IFNs and TLR signaling. While NK cells, DC, especially pDC, as well as macrophages and other innate factors, are required to eradicate both HSV-1 and HSV-2, it is well established that adaptive immune responses, especially CD4+ Th1 cells except CD8+ T cells, are necessary to clear the infection. Although functional adaptive immunity is required to clear viral infection, these responses alone are not sufficient to protect against infection in the absence of innate immune mechanisms. Therefore, many studies investigate the protective effect and immune modulatory role of drugs, natural products such as plant extracts and prophylactics agents against mucosal herpes infection. Collectively, it is important to consider a functional innate defense and its role in assisting subsequent adaptive immune responses, when developing effective therapeutic and vaccine strategies against both HSV-1 and HSV-2.
Go to :
 XML Download
XML Download