

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
DC
dendritic cell
cDC
conventional dendritic cell
pDC
plasmacytoid dendritic cell
moDC
monocyte-derived dendritic cell
Treg
regulatory T cells
PRRs
pattern recognition receptors
TLR
toll-like receptor
DTR
diphtheria toxin receptor
IFN
interferon
Tip-DC
TNF and iNOS derived NO producing dendritic cell
INTRODUCTION
The lung is the essential organ for respiration. Because the lung mucosal area contacts air for gas exchange, it can be infected easily by various microbes, such as influenza, respiratory syncytial virus (RSV), pneumococcus, and Aspergillus. Nevertheless, the lung possesses a sentinel system that identifies these threats and elicits an anti-microbial response. In this review, we focus on the immune response to respiratory viral infection, which can induce acute respiratory disease.
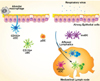
Dendritic cells (DCs) participate in the first line of defense in the innate immune response against respiratory viral infection. DCs are distributed throughout the entire lung, with each subset localized to a specific compartment of the organ (1). In the absence of inflammation, lung DCs can be subdivided into three distinct subsets based on the combined expression of cell surface markers: CD103+ conventional DCs (cDCs), CD11b+ cDCs, and plasmacytoid DCs (pDCs). During inflammation, monocyte-derived DCs (moDCs) are generated in the lung (2,3) (Table I).
In a respiratory virus infection, one virus can induce different types of immune responses depending on the type of DC subset activated (4,5). In this process, cell type-specific pattern recognition receptors (PRRs) may also be involved (6). Each DC subset expresses different pattern recognition receptors, thereby enabling the cells to react differently depending on the type of virus infection (7). In particular, neither a vaccine nor an effective antiviral therapy is currently available against RSV infection (8). To development vaccine for RSV infection, understanding the role of the lung DC subsets is important. Determining the specialized functions of the various lung DC subsets is challenging. This review focuses on the distinctive features and antiviral functions exhibited by the various lung DC subsets during respiratory virus infection in mice.
CD103+ conventional dendritic cells
The CD103+CD11b- cDC subset shares its origin and function with lymphoid tissue CD8α+ cDCs (9,10). CD103+ cDCs are primarily distributed to connective tissues. The proportion of CD103+ cDCs among total conventional DCs rarely exceeds 20~30%. These cells express higher fms-like tyrosine kinase 3 (Flt3) levels compared to CD11b+ cDCs and therefore proliferate in response to Flt3 ligand (11). CD103 expression is dependent on the tissue microenvironment and regulated by local production of the cytokine Csf-2 (GM-CSF) (12,13,14,15). However, CD103-deficient mice do not exhibit major defects in DC development (16). CD103+ cDCs lack the macrophage-related markers CD11b, CD115, CD172a, F4/80, and CX3CR1. With the exception of intestinal and pancreatic CD103+ cDCs, these cells express the C-type lectin receptor langerin (11,17).
Besides connective tissues, CD103+ cDCs are located in nonlymphoid tissues at the interface with the environment. Lung CD103+ cDCs can be found in the mucosa and vascular wall (18) (Fig. 1). Following antigen uptake, CD103+ cDCs migrate to the T cell zone of draining lymph nodes (10). In the airways and gut, DCs extend their processes between epithelial cells to contact the airway lumen directly. These airway mucosal DCs can conduct continuous immune surveillance of the airway luminal surface, thereby acting like a periscope (19,20,21). In mouse lungs, intraepithelial CD103+ cDCs express the tight-junction proteins claudin-1, claudin-7, and zonula-2, which form tight junctions with airway epithelial cells (18). As a result, CD103+ cDCs can sample contents within the airway lumen without disturbing the function of the epithelium barrier.
Current reports have shown that, following influenza or RSV infection, CD103+ cDCs migrate from the intraepithelial base to the draining mediastinal lymph nodes (22,23), where they mainly present antigen to naïve CD4+ and CD8+ T cells. However, previous studies demonstrated that antigens are transferred from migratory DCs to CD8α+ resident cDCs, and presented to T cells by antigen-bearing CD8α+ resident cDCs (24).
As previously mentioned, studies have established that CD103+ cDCs belong to the CD8α+ subset of cDCs (2). Like lymphoid-derived CD8α+ cDCs, CD103+ cDCs originate exclusively from pre-DCs under the control of Flt3 ligand, inhibitor of DNA protein 2 (Id2), and interferon regulatory protein 8 (Irf8) (11,25). Murphy et al. reported a developmental relationship between lymphoid organ-resident CD8α+ cDCs and nonlymphoid CD103+ cDCs using Batf3-deficient mice (26). A recent study used heat maps to demonstrate that the expression of pattern recognition receptors, cytokines, and chemokine receptors is similar between CD103+ cDCs and CD8α+ cDCs (27). In particular, both subsets express TLR3, TLR11, the scavenger receptor CD36, and C-type lectin Clec9A (28,29,30,31). Desch, et al. showed that mouse lung CD103+ cDCs selectively express TLR3, while CD11b+ cDCs express TLR2 and TLR7 (32).
CD103+ cDCs play a nonredundant role in stimulating CD8+ T cell-mediated immunity. Influenza virus infection following depletion of CD103+ cDCs in langerin-diphtheria toxin receptor (langerin-DTR) mice results in severe illness, defective viral clearance, and abrogated antiviral response due to impaired development of influenza virus-specific CD8+ T cells (22). In the Batf3 knockout mouse model, the CD103+ cDC-deficient mice cannot produce CD8+ T cell priming in response to influenza infection (33). CD103+ cDCs play an important role in cross-presentation of apoptotic cell-associated antigen to CD8+ T cells (25,32). However, whether CD103+ cDCs can induce a cytotoxic T cell response against RSV infection (similar to other viruses) remains to be investigated.
The role of lung CD103+ cDCs in the activation of CD4+ T cells is unclear. In cutaneous skin infection with Candida albicans, dermal CD103+ cDCs control the induction of pathogen-specific CD4+ IFN-γ+ T cells (34). A recent study using langerin-DTR mice demonstrated that ablation of CD103+ cDCs inhibited induction of the encephalitogenic CD4+ Th1 response and autoimmune encephalomyelitis (EAE) (35). However, some studies showed that the CD4+ T cell response was independent of CD103+ cDCs. Batf3 knockout mice that are deficient in CD103+ cDCs can mount an efficient CD4+ T cell response to West Nile virus or autoimmune EAE (14,25). Moreover, ablation of CD103+ cDCs in langerin-DTR mice did not affect the CD4+ T cell response against Leishmania major infection (36).
CD8α+ cDCs and CD103+ cDCs are thought to participate in deletional tolerance of self-reactive T cells and the induction of antigen-specific regulatory T cells (Treg) (16). Splenic DCs captured dying cells and processed, then induced specific tolerance (37,38). A report showed that the CD103+CD207+ subset of splenic CD8α+ cDCs is responsible for tolerance induction to cell-associated antigens (39). However, an autoimmune response was not observed in Batf3 knockout mice that lack CD8α+ cDCs and CD103+ cDCs. Thus, the tolerogenic function of lung CD103+ cDCs remains to be determined.
CD11b+ conventional dendritic cells
In the lung, CD11b+ cDCs reside mainly in the lamina propria, which is located below the basement membrane (Fig. 1). CD11b+ cDCs are heterogeneous and their development depends on both Flt3 and M-CSFR (11). Dependency on M-CSFR is suggestive of a monocytic origin, and some non-lymphoid CD11b+ cDCs can be reconstituted by pre-DC. CD11b+ cDCs frequently lack CD103 but express CD11b. Despite this, markers to distinguish the two ontogenically distinct subsets differ between tissues. For instance, expression of CD64 (FcγRI) helps distinguish between these two subpopulations in muscle, whereas expression of CD103 helps discriminate between the two CD11b+ DC subsets in the intestinal lamina propria (40,41). Lambrecht et al. recommended detection of CD64 and MAR-1 expression as the most reliable method to discriminate between monocyte-derived DCs and CD11b+ cDCs in the lung and mediastinal lymph nodes (42).
Because CD11b+ cDCs are not a homogenous subset, the exact PRR profile of CD11b+ cDCs is complex. Nevertheless, these receptors are expressed differentially in CD103+ cDCs and CD8α+ cDCs (27). Quantitative proteomics has revealed that splenic CD11b+ cDCs express high levels of cytoplasmic viral sensors and are potent cytokine producers in the steady state and upon stimulation (43). Lung CD11b+ cDCs are major producers of proinflammatory chemokines, including MCP-1, MIP-1α, MIP-1β, RANTES, and MCP5, attracting inflammatory cells and effector T lymphocytes to the lung (44).
CD11b+ cDCs can capture antigens and migrate from nonlymphoid tissues to regional draining lymph nodes (23). Research has established that CD8α+ cDCs and CD103+ cDCs play crucial roles in cross-presentation. However, during influenza infection, CD103+ cDCs and CD11b+ cDCs are the primary mediators of antigen presentation to naïve CD8+ T cells in the draining lymph nodes (45).
During severe influenza infection, CD11b+ cDCs, but not CD103+ cDCs or CD8α+ resident cDCs, accumulate in the draining lymph nodes to become the predominant DC subset responsible for stimulating CD8+ T cells via the costimulatory molecule CD70 (46). These contradictory findings could be attributed to the different viral doses used for infection and the differential effects of direct DC infection by influenza virus. Severe viral infection induced CD11b+ cDCs that were incapable of antigen presentation to CD8+ T cells. However, low viral doses enabled directly infected CD11b+ cDCs to arrive at the draining lymph nodes ready to prime the CD8+ T cell response (47). In addition, CD11b+ cDCs are thought to play a predominant role in MHC class II presentation, including acting as the predominant presenters of viral antigens to CD4+ T cells in response to influenza virus infection (45).
CD11b+ cDCs constantly escape from the blood to the thymus to induce central tolerance, such as clonal deletion of autoreactive T cells or differentiation of Treg (48,49). CD103+ CD11b+ cDCs purified from the lamina propria of the small intestine were found to promote a high level of Treg differentiation relative to lymphoid organ-derived DCs (50,51). However, the contribution of lung CD11b+ cDCs in tolerance has not been established.
In addition to CD103+ cDC-mediated uptake in the airways, CD11b+ cDCs utilize another pathway to acquire inhaled antigens. TLR4 triggering of epithelial cells caused production of innate proallergic cytokines, including thymic stromal lymphopoietin (TSLP), granulocyte-macrophage colony-stimulating factor (GM-CSF), interleukin-25, and interleukin-33. In the absence of TLR4 on structural, but not hematopoietic cells, CD11b+ cDCs were not recruited or activated in a chimeric mouse model (52). It is unclear whether lung CD11b+ cDCs require epithelial activation as well.
CD11b+ cDCs are essential for the maintenance of inducible bronchus-associated lymphoid tissue (iBALT), a tertiary lymphoid organ (TLO) induced in the lungs after influenza infection (53). After viral clearance, CD11b+ cDCs isolated from the lungs of mice with iBALT no longer presented viral antigens to T cells but produced lymphotoxin (LT) β and homeostatic chemokines (CXCL-12, CXCL-13, CCL-19, and CCL-21) known to contribute to TLO organization. Using the replication-deficient modified vaccinia virus model, Halle, et al. described iBALT as a tertiary lymphoid structure that supports the efficient priming of T cells against unrelated inhaled antigens with DCs required for its maintenance (54).
Plasmacytoid dendritic cells
pDCs are distributed to conducting airways as well as parenchyma and alveolar septa in the lung (Fig. 1). These cells represent a small subset of DCs, which share a common origin with cDCs. pDCs develop in the bone marrow from a continuum of Flt3+c-Kitlow progenitors, including lymphoid progenitors and common DC progenitors (CDPs). Their development proceeds through the putative committed pDC progenitor and immature pDCs in the bone marrow toward the mature peripheral pDCs (55). Upregulation of the basic helix-loop-helix transcription factor (E protein) E2-2 serves as a key lineage commitment event in pDC development (56,57). Because E proteins are essential regulators of lymphocyte development, E2-2 activity may underlie the distinct lymphoid features of pDCs. These cells express low levels of MHC class II and costimulatory molecules, as well as low levels of CD11c in the steady state (16). They also express a narrow range of PRRs, including TLR7 and TLR9.
Generally, pDCs function during the antiviral response to produce type I IFNs that induce the adaptive immune response. Some studies have shown that pDCs can trigger an influenza-specific CD8+ T cell response in vitro (58,59,60). However, RSV-stimulated pDCs cannot enhance the proliferation and maturation of antigen-specific T cells, but rather promote direct antiviral activity by secreting type I IFNs (61).
Following influenza infection in vivo, 120G8+CD11cint pDCs accumulate in the lung and lymph nodes carrying viral nucleoprotein (NP). Depletion of pDCs using 120G8 antibodies did not affect viral clearance or clinical severity during influenza infection (22). Instead, pDC depletion led to a reduction in antiviral antibody production after clearance of influenza from the lung. However, depletion of pDCs resulted in decreases viral clearance of RSV infection and exacerbation of all facets of immune-mediated pathology, including increase of airway hyper-responsiveness, pulmonary inflammation, and mucus production (62,63).
In IkarosL/L mice, expressing low levels of the transcription factor Ikaros (Ik(L/L)) lack peripheral pDCs, pDCs regulate T cell accumulation in the bronchoalveolar space during early influenza virus infection, but are not essential for controlling this disease (64). These data demonstrated that the antiviral CD8+ T cell response was independent of pDCs. However, in BDCA2-DTR mice, pDC depletion reduced early type I IFN production, enhanced early viral replication, and impaired the survival and accumulation of virus-specific cytotoxic T lymphocytes in systemic MCMV or VSV infection (65).
According to a recent report, pDCs do not appear to influence viral burden, survival, or virus-specific CD8+ T cell response during local HSV infection. In contrast, pDCs were important for early type I IFN production, NK cell activation, and CD8+ T cell response during systemic HSV infection (66). These results help elucidate the antiviral role of pDCs in respiratory virus infection. However, whether pDCs can differentially respond under different conditions between host and virus remains to be determined.
Monocyte-derived dendritic cells
Inflammatory moDCs differentiate from circulating Ly6Chi monocytes (67) (Fig. 1). Recent studies have established that, under conditions of stress, such as TLR stimulation, early hematopoietic precursors can differentiate into DCs, bypassing normal growth and differentiation requirements (68,69). However, the contribution of monocytes and DC-related precursors to the differentiation of lung moDCs in response to respiratory virus infection remains unclear.
Most inflammatory DCs are characterized by the expression of Ly6C, CD11b, MHC class II, and intermediate levels of CD11c (67). Ly6C is a distinct marker of monocytes, but that is downregulated rapidly in the presence of moDCs (42,70,71). Therefore, distinguishing inflammatory moDCs from nonlymphoid CD11b+ DCs is challenging. As mentioned in the preceding section, one report demonstrated that staining with the MAR-1 antibody directed against the high affinity immunoglobulin E (IgE) α chain receptor (FcεRI) is better than staining for Ly6C (2). A recent study showed that inflammatory moDCs are recruited to draining lymph nodes following lipopolysaccharide (LPS) stimulation, and that these moDCs express the lectin DC-SIGN/CD209, the mannose receptor CD206, and CD14 (71).
Monocytes were originally considered the immediate upstream precursors of cDCs. This hypothesis originated from studies showing that DCs could be differentiated in vitro from human blood mononuclear cells using GM-CSF and IL-4 (72). When monocytes were transferred into mice with an inflammatory milieu dependent on GM-CSF, monocytes produced a distinct type of splenic DC (73). Nowadays, the concept that monocytes are a precursor of inflammatory DCs is widely accepted. More recent studies have shown that monocytes contribute to cDC development in the steady state (41,74,75,76). However, because this review focuses on DC subsets that act against respiratory virus infection, we refer to mononuclear cell-derived DCs as moDCs in inflammation.
CD11b+ DCs can produce TNF and iNOS-derived NO during L. monocytogenes infection. These Tip-DCs are dependent on CCR2 and mediate innate immunity against this intracellular bacterial pathogen (77), suggesting that Tip-DCs may contribute to the elimination of intracellular pathogens.
A recent study identified an uncharacterized zinc finger transcription factor named zDC (Zbtb46, Btbd4) that is expressed specifically by cDCs and committed cDC precursors but not by monocytes, pDCs, or other immune cell populations (78,79). zDC-DTR mice treated with diphtheria toxin eliminated LPS-induced inflammatory moDCs, suggesting that LPS induced inflammatory moDCs that belong to a real DC population. However, L. monocytogenes infection-induced Tip-DCs were not ablated by DT treatment in these mice. Given this result, Tip-DCs most likely resemble monocytes more than DCs.
CD11b+ moDCs are recruited to inflammatory sites in the lungs following exposure to respiratory antigen or virus. During influenza infection, moDCs also differentiate from monocytes in the lung. These trafficking and differentiation process are dependent on type I IFN signaling and CCR2 during influenza infection (80,81). Some in vitro studies suggested that type I IFN-producing moDCs can regulate viral replication (82,83); however, whether moDCs participate directly in the antiviral response remains unclear. Interestingly, CCR2-deficient mice did not exhibit increased influenza viral titer.
Whether moDCs can migrate to draining lymph nodes and induce the T cell response has not been determined (45,46). Monocyte-derived CD11c+ DCs, which express CX3CR1, can patrol the vessel wall of the pulmonary arterial vasculature and capture embolic materials. Thus, these cells are essential and sufficient for priming of naïve T cells in lung draining mediastinal lymph nodes (84). Some studies have shown that moDCs may be important for the interaction of effector T cells present in the infection site instead of the lymph nodes (85,86).
CD8α+ dendritic cells
Generally, CD8α+ cDCs do not exist in the lung because these cells are non-migrating, lymphoid-organ resident DCs. However, CD8α+ cDCs are involved in respiratory virus infection. They can induce the T cell response in mediastinal lymph nodes. CD8α+ cDCs constitute 20~40% of spleen and lymph node cDCs. Similar to CD103+ cDCs, CD8α+ cDCs lack expression of CD11b and other macrophage markers. However, they express high levels of Flt3 and proliferate in response to Flt3 ligand (87).
Lymphoid resident CD8α+ cDCs are immature in the steady state, but microbial products can induce maturation of CD8α+ cDCs. Lymph node CD8α+ cDCs are located in the subcapsular sinus, the site of afferent lymphatic vessel entry (39,88). After antigen uptake, these CD8α+ cDCs migrate to the T cell zone where they present antigens.
As mentioned above, CD8α+ cDCs share their origin and function with nonlymphoid CD103+ cDCs. However, the function of CD8α+ cDCs themselves is still unclear. Generation of conditional or knockout mouse models for specific depletion of CD8α+ cDCs will aid in our understanding of the function of these cells. Additional studies are required to determine whether these cells have a common immediate precursor and to investigate which cell is the precursor and progeny for CD8α+ cDC (89).
CONCLUSION
Respiratory viruses can induce acute respiratory disease. In the lung, DCs are the first line of sentinel cells in the innate immune response against respiratory viral infection, similar to alveolar macrophages. DCs are crucial in regulating the immune response by bridging innate and adaptive immunity. These cells can produce inflammatory cytokines and chemokines, as well as migrate to the draining lymph nodes to initiate the adaptive immune response through antigen presentation. Lung DCs associated with viral infection can be subdivided into CD103+ cDCs, CD11b+ cDCs, pDCs, and moDCs. Lymphoid CD8α+ cDCs also play an important role in the antiviral response. These different DC subsets have functional specialization against respiratory viral infection. One virus can induce different immune responses depending on the type of DC subset activated. Moreover, one subset can react differently depending on the type of virus encountered. Contextually controlling the balance between these specialized DC subsets is important for an effective antiviral response and maintaining immune homeostasis. Moreover, understanding the differential roles of lung dendritic cell subsets against respiratory virus infection is a key point to develop a vaccine.

 XML Download
XML Download