

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Immune responses against self-antigens caused by failure of self-tolerance develop autoimmune diseases which affect approximately 7~9% of the general population (1). However, current treatments of autoimmune diseases are not curative and induce general immunosuppression which is associated with serious infection and other adverse outcomes (2). Therefore, it is necessary to develop new therapeutics in order to selectively suppress autoreactive immune cells, including B cells, CD4+ T cells, and CD8+ T cells.
Unlike B cells, T cells recognize epitope peptides presented by major histocompatibility complex (MHC) molecules. Binding of T-cell epitope peptides to MHC is determined by hydrophobic or charge interactions between the residue of peptides and the cleft of MHC molecules (3,4). Thus, peptides presented on MHC molecules can be replaced extracellularly by other peptides depending on their affinity to MHC molecules (5,6). If peptides with high affinity to MHC molecules are present extracellularly in excess, they would occupy the cleft of MHC molecules and replace preexisting peptides on MHC molecules. This concept might be exploited for the treatment of autoimmune diseases. Autoimmune responses induced by self peptide-specific T cells could be inhibited by the addition of competitive peptides that replace self peptides on MHC molecules and inhibit presentation of the self peptide on antigen presenting cells (APCs).
In the present study, we investigated if priming of autoreactive CD8+ T cells would be inhibited by competitive peptides for MHC class I binding. We used a mouse model of vitiligo which is induced by immunization of Kb-binding tyrosinase-related protein 2 (TRP2)-180 peptide. Previously, we established a novel mouse model of vitiligo by immunization with TRP2-180 peptide in combination with lipopolysaccharides (LPS) and CpG oligodeoxynucleotides (CpG ODN) (7). This mouse model of vitiligo was characterized by progressive depigmentation of the skin due to selective loss of melanocytes. In addition, the extent of skin depigmentation correlated with the frequency of TRP2-180-specific CD8+ effector T cells in the spleen. Therefore, we used this mouse model of vitiligo to analyze how efficiently the competitive peptides for MHC class I binding inhibit priming and activation of melanocyte-specific autoreactive CD8+ T cells.
In this study, competitive peptides for Kb binding inhibited IFN-γ production and proliferation of TRP2-180-specific CD8+ T cells upon ex vivo peptide restimulation. Interestingly, the capability of inhibition was influenced by T-cell immunogenicity of the competitive peptides in vivo. The competitive peptide with a high T-cell immunogenicity efficiently inhibited priming of TRP2-180-specific CD8+ T cells in vivo, whereas the competitive peptide with a low T-cell immunogenicity did not. Taken together, the inhibition of the priming of autoreactive CD8+ T cells depends on not only competition of peptides for MHC class I binding but also competitive peptide-specific CD8+ T cells. Thus, highly T cell-immunogenic peptides which are competitive for MHC binding might be useful for inhibiting priming of autoreactive T cells for the treatment of autoimmune diseases.
MATERIALS AND METHODS
Animals and peptides
Male C57BL/6 mice were kept in accordance with the institutional guidelines, and used at the age of 4~5 weeks for peptide immunization. Peptides were purchased from Peptron (Daejeon, Korea) and dissolved in 5% DMSO phosphate-buffered solution (PBS).
Measurement of relative Kb binding affinity of peptides
Splenocytes of naive C57BL/6 were incubated with different combinations of peptides, including SIINFEKL at 0.5µM and various concentrations of additional peptides (0.5, 5, or 50µM) as indicated, at 37℃ for 4 hrs. After incubation, splenocytes were washed and fixed by 4% paraformaldehyde. The density of SIINFEKL-binding Kb molecules on lymphocytes was analyzed by flow cytometry using 25-D1.16 monoclonal antibody (8) that reacts with SIINFEKL-binding Kb molecule. The percentage of decreased density of SIINFEKL-binding Kb molecules by additional peptides was calculated as follows: [100-mean fluorescence index (MFI) of SIINFEKL-binding Kb in SIINFEKL at 0.5µM with additional peptides at 5µM/MFI of SIINFEKL-binding Kb in SIINFEKL at 0.5µM×100].
Inhibition of IFN-γ production by competitive peptides in vitro
C57BL/6 mice were subcutaneously immunized twice at 1-week intervals in the footpad with TRP2-180 (50µg), LPS (5µg), and CpG ODN (5µg). One week after the second immunization, splenocytes were harvested and stimulated with different combinations of peptides, including TRP2-180 at 0.1µM and various doses of additional peptides (0, 0.1, 1, or 10µM). TRP2-180-specific IFN-γ production was analyzed by flow cytometry after intracellular cytokine staining (ICS) as described in the section of 'Intracellulr Cytokine Staining'.
Generation of TRP2-180-specific CD8+ T cell lines
CD45.2 C57BL/6 mice were subcutaneously immunized 3 times at 1-week intervals in the hind footpad with TRP2-180 (50µg), LPS (5µg) and CpG ODN (5µg). One week after the last immunization, splenocytes were harvested, and splenic CD8+ T cells were sorted using anti-CD8 microbeads (Miltenyi Biotec, Auburn, CA). These sorted CD8+ T cells were restimulated in vitro 3 times at 2-week intervals with TRP2-180 peptide (10µg/ml) in the presence of IL-2 (20 IU/ml) and CD45.1 splenocytes as APCs. The purity of TRP2-180-specific CD8+ T cells was evaluated by TRP2-180-specific IFN-γ production and was >80%. Two weeks after the last stimulation, CD8+ T cells were used for proliferation analysis.
Inhibition of T cells proliferation by competitive peptides in vitro
CD45.2+, TRP2-180-specific CD8+ T cell line (0.2×106 cells) labeled with CFSE at 10µM (Invitrogen Ltd, Paisley, UK) was cocultured with CD45.1+ splenocytes (1×106 cells) pulsed with different combinations of peptides, including TRP2-180 at 0.1µM and additional peptides at 10µM. Four days after coculture, TRP2-180-specifc proliferation of CD8+ T cells was evaluated by fluorescence intensity of CFSE in flow cytometry analysis. TRP2-180-specific CD8+ T cells were distinguished from antigen presenting splenocytes by CD45.2 gating (anti-CD45.2-APC-H7, BD Biosciences, San Jose, CA, USA), and dead cells were excluded using 7-Amino-Acinomycin D (7-AAD, BD Biosciences). The percentage of proliferation inhibition by additional peptides was calculated as follows: [100-(% of proliferation in TRP2-180 at 0.1µM with additional peptides-% of proliferation in the absence of TRP2-180)/(% of proliferation in TRP2-180 at 0.1µM-% of proliferation in the absence of TRP2-180)×100].
Inhibition of priming of TRP2-180-specific CD8+ T cells in vivo
C57BL/6 mice were subcutaneously immunized in the footpad with TRP2-180 (5 nmole), LPS (5µg), and CpG ODN (5µg). One day after the immunization, additional competitive peptides (50 nmole) were injected into the same footpad without adjuvants twice at 1-day intervals. One week after TRP2-180 immunization, splenocytes were harvested and stimulated with TRP-180 or competitive peptides, and peptide-specific IFN-γ+ CD8+ T cell population was analyzed by flow cytometry after ICS.
Evaluation of disease progression in a vitiligo mouse model
Vitiligo was induced by TRP2-180 immunization in mice as described previously (7). Briefly, C57BL/6 mice were subcutaneously immunized in the footpad with TRP2-180 (5 nmole), LPS (5µg) and CpG ODN (5µg) twice at 1-week intervals. One week after the second immunization, TRP2-180 (5 nmole) with the same adjuvants was injected into the tail dermis for disease induction twice at 1-week intervals. In this model, disease progression was evaluated with or without administration of competitive peptides. Competitive peptides (50 nmole) were injected into the tail dermis 1 day after third and fourth TRP2-180 immunization without adjuvants twice at 1-day intervals. Five weeks after the last TRP2-180 immunization, splenocytes were harvested and stimulated with TRP-180 or competitive peptides. Peptide-specific IFN-γ+ CD8+ T-cell population was analyzed by flow cytometry after ICS. In addition, the area of depigmented skin per tail was calculated by using the ImageJ program (National Institutes of Health, Bethesda, MD, USA) and presented as the percentage of depigmented skin per tail.
Intracellular Cytokine Staining (ICS)
Splenocytes were resuspended at 107 cells/ml in RPMI 1640 containing 1% FBS, and 200µl of the suspension was added per well. Single cell suspensions were stimulated with TRP2-180 (5µg/ml) or additional competitive peptides (5µg/ml), and brefeldin A (GolgiPlug, BD Biosciences) was added 1 hr later. After another 5 hrs of incubation at 37℃, cells were stained with ethidium monoazide (Sigma-Aldrich, St. Louis, MO, USA). After washing, the cells were stained with anti-CD3-pacific blue, anti-CD4-APC-Cy7, anti-CD8-PE-Cy7, and anti-CD44-PE (all from BD Biosciences), permeabilized using a Cytofix/Cytoperm kit (BD Biosciences), and stained with anti-IFN-γ-APC (BD Biosciences). Stained cells were analyzed on an LSR flow cytometer (BD Biosciences).
RESULTS
Relative affinity of peptides for Kb binding
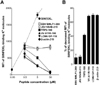
In the present study, Kb-binding TRP2-180 peptide was used as a self peptide to induce melanocyte-specific CD8+ T cells and develop vitiligo (Table I) (7,9). For competition with TRP2-180 for Kb binding, we included vaccinia virus A11R 198-205 (VV A11R-198) (10), lymphocytic choriomeningitis virus GP 118-125 (LCMV GP-118) (10), and β-actin 219-227 (β-actin-219) (11) which bind to Kb. We also included Japanese encephalitis virus NS4B 215-223 (JEV NS4B-215) (12) which bind to Db, and Epstein-Barr virus BMLF1 280-288 (EBV BMLF1-280) (13) which bind to human leukocyte antigen (HLA)-A0201, as non-competitive peptides for Kb binding.
First, we evaluated relative Kb-binding affinity of the peptides by a competition assay using SIINFEKL peptide. In this assay, splenocytes were pulsed with SIINFEKL peptide in combination with competitive peptides, then the density of SIINFEKL-binding Kb molecules was quantified by flow cytometry using antibody specific to SIINFEKL-binding Kb molecule. Without competitive peptides, SIINFEKL peptide pulsing increased the density of SIINFEKL-binding Kb molecule in a dose-dependent manner (Fig. 1A). When splenocytes were pulsed with a fixed concentration (0.5µM) of SIINFEKL peptide in combination with increased concentration of additional peptides, Kb-binding peptides (TRP2-180, VV A11R-198, LCMV GP-118, and β-actin-219) decreased the density of SIINFEKL-binding Kb molecules in a dose-dependent manner (Fig. 1A). In particular, VV A11R-198, LCMV GP-118, and β-actin-219 more decreased the density of SIINFEKL-binding Kb molecules than TRP2-180 (Fig. 1A), indicating that they have a higher affinity for Kb than TRP2-180. Non-Kb-binding peptides (JEV NS4B-215 and EBV BMLF1-280) did not decrease the density of SIINFEKL-binding Kb molecules (Fig. 1A), confirming that they do not bind to Kb. By calculating the percentage of decreased density of SIINFEKL-binding Kb molecules, the relative Kb-binding affinity of peptide was determined (Fig. 1B).
Inhibition of TRP2-180-specific IFN-γ production by competitive peptides in vitro
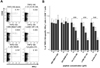
To evaluate the inhibition of TRP2-180-specific IFN-γ production by competitive peptides, splenocytes of TRP2-180-immunized mice were stimulated ex vivo with TRP2-180 peptide alone or in combination with competitive peptides, and IFN-γ ICS was performed. IFN-γ production was observed in CD44+, antigen-experienced CD8+ T cells. As expected from the relative Kb-binding affinity of each peptide, VV A11R-198, LCMV GP-118, and β-actin-219 significantly inhibited IFN-γ production of TRP2-180-specific T cells in a dose-dependent manner, while JEV NS4B-215 and EBV BMLF1-280 did not (Fig. 2A and B).
Inhibition of TRP2-180-specific T-cell proliferation by competitive peptides in vitro
To examine the inhibition of T cells proliferation by competitive peptides in vitro, a TRP2-180-specific T-cell line was established, and their proliferation responding to TRP2-180 peptide was evaluated by CFSE labeling. Proliferation of the T-cell line was increased by stimulation with TRP2-180 peptide (Fig. 3A). VV A11R-198, LCMV GP-118, and β-actin-219 inhibited TRP2-180-specific proliferation of T cells, while JEV NS4B-215 and EBV BMLF1-280 did not (Fig. 3B). The percentage of proliferation inhibition was calculated and presented for each peptide (Fig. 3C).
Inhibition of TRP2-180-specific T-cell priming by competitive peptides in vivo
Next, we determined if Kb-binding competitive peptides would inhibit priming of TRP2-180-specific T cells in vivo. C57BL/6 mice were subcutaneously immunized in the footpad with TRP2-180, LPS, and CpG ODN. One day after the immunization, doses 10-times higher than competitive peptides were injected into the same footpad without adjuvants twice at 1-day intervals. One week after TRP2-180 immunization, splenocytes were harvested and TRP2-180-specific IFN-γ+ CD8+ T cells were analyzed by ICS and flow cytometry. Competitive peptide-specific IFN-γ+ CD8+ T cells were also analyzed. Non-Kb-binding peptides (JEV NS4B-215 and EBV BMLF1-280) did not decrease the frequency of TRP2-180-specific IFN-γ+ CD8+ T cells as expected (Fig. 4A). Among the Kb-binding peptides, only VV A11R-198 significantly decreased the frequency of TRP2-180-specific IFN-γ+ CD8+ T cells (Fig. 4A). LCMV GP-118 showed a tendency to decrease the frequency of TRP2-180-specific IFN-γ+ CD8+ T cells (Fig. 4A), while β-actin-219 did not (Fig. 4A). Interestingly, the ability of Kb-binding peptides to inhibit TRP2-180-specific T-cell priming was associated with T-cell immunogenicity of each peptide. VV A11R-198 which significantly inhibited TRP2-180-specific T-cell priming showed the highest T-cell immunogenicity (Fig. 4B), and β-actin-219 which did not inhibit the priming did not show T-cell immunogenicity (Fig. 4B). The data suggest that inhibition of priming of autoreactive CD8+ T cells depends on not only the competition of peptides for MHC class I binding but also competitive peptide-specific CD8+ T cells.
Competitive peptides do not inhibit disease progression in a mouse model of vitiligo
C57BL/6 mice were subcutaneously immunized in the footpad with TRP2-180, LPS, and CpG ODN twice at 1-week intervals. One week after the second immunization, TRP2-180 with the same adjuvants was injected into the tail dermis for disease induction twice at 1-week intervals. Competitive peptides were injected into the tail dermis 1 day after the third and fourth TRP2-180 immunization without adjuvants twice at 1-day intervals. Five weeks after the last TRP2-180 immunization, splenocytes were harvested and TRP2-180-specific IFN-γ+ CD8+ T cells were analyzed by ICS and flow cytometry. Competitive peptide-specific IFN-γ+ CD8+ T cells were also analyzed. In this case, no peptide decreased the frequency of TRP2-180-specific IFN-γ+ CD8+ T cells, while the 2 Kb-binding peptides (VV A11R-198 and LCMV GP-118) were immunogenic (Fig. 5A and B). As expected from the fact that no peptide decreased the frequency of TRP2-180-specific IFN-γ+ CD8+ T cells, disease progression evaluated by the percentage of depigmented skin lesions in a tail was not prevented by any peptide (Fig. 5C). VV A11R-198 showed a tendency to decrease the area of depigmented lesions in a tail, without statistical significance (Fig. 5C).
DISCUSSION
In the present study, we employed competitive peptides for Kb binding to inhibit activation of TRP2-180-specific CD8+ T cells, melanocyte-specific autoreactive CD8+ T cells. The competitive peptides displayed a higher Kb-binding affinity than TRP2-180 peptide, an autoimmunogenic peptide (Fig. 1A and B) and inhibited IFN-γ production and proliferation of TRP2-180-specific CD8+ T cells in vitro, while non-Kb-binding peptides did not (Fig. 2 and 3). The data demonstrate that the inhibition of TRP2-180-specific CD8+ T-cell activation in vitro depends on competition of peptides for MHC binding.
When we tested if Kb-binding competitive peptides inhibit priming of TRP2-180-specific CD8+ T cells, only VV A11R-198 significantly inhibited the priming (Fig. 4A), while β-actin-219 did not (Fig. 4A). Interestingly, the ability of Kb-binding peptides to inhibit TRP2-180-specific T-cell priming was associated with T-cell immunogenicity of each peptide. In fact, VV A11R-198 showed the highest immunogenicity to T cells (Fig. 4B), and β-actin-219 did not show any immunogenicity (Fig. 4B). These results suggest that clonal expansion of autoreactive T cells might be affected by expansion of competitive peptide-specific T cells. These findings are reminiscent of clonal competition in immunodominance and immune hierarchy in CD8+ T-cell responses (14). T cells primed by highly immunogenic peptides compete with autoreactive T cells for resources required for T-cell survival and proliferation (15). Therefore, we should consider not only competition of therapeutic peptides for MHC class I binding but also T-cell immunogenicity of therapeutic peptides in competitive peptides-based therapy for the treatment of autoimmune diseases.
Although VV A11R-198 inhibited the priming of TRP2-180-specific CD8+ T cells (Fig. 4A), it did not inhibit the boosting of them (Fig. 5A), and thus disease progression was not prevented by VV A11R-198 in a mouse model of vitiligo (Fig. 5C). The results demonstrate that VV A11R-198 could inhibit priming of naive T cells in vivo but could not inhibit reactivation of memory T cells in the tail skin. Since memory T cells rapidly proliferated and efficiently differentiated to effector T cells compared with naive T cells (16,17), our results could be explained by the difference between memory T cells and naive T cells. If competitive therapeutic peptides also target memory T cells, autoreactive memory T cells might be inhibited by competitive peptides. In this context, T-cell epitope peptides from influenza A virus or cytomegalovirus will be candidates for competitive peptides-based therapy because the majority of human population have a memory T-cell pool against these viruses (18,19).
In summary, competitive peptides for Kb binding inhibited IFN-γ production and proliferation of TRP2-180-specific CD8+ T cell in vitro. In mice, inhibition of TRP2-180-specific CD8+ T-cell priming required competitive peptides with a high immunogenicity for T cells. Although competitive peptides did not prevent the progression of the depigmented skin lesion in a mouse model of vitiligo, our results provide new insights into the development of competitive peptides-based therapy for the treatment of autoimmune diseases.




 XML Download
XML Download