

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Basophils are considered one of the major cell types associated with Th2 immunity (1-4). Basophils exert immunomodulatory roles mainly by producing IL-4 and by functioning as professional Ag presenting cells (3-5), although the latter possibility was recently challenged by others (6-9). It was recently proposed that a subset of dendritic cells (DC) expressing IgE receptor (FcεRI) is highly specialized in generating Th2 type effector cells (7,9). In addition, recent identification of innate type 2 cells and Th2-inducing cytokines adds additional complexity to in vivo Th2 immunity. For example, epithelial cell-derived cytokines such as IL-25 induces Th2 immune responses especially at the mucosal surface following helminth infections (10,11). IL-25 also promotes accumulation of multi-potent progenitor (MPP) type 2 cells in the gut associated lymphoid tissues (12). Adoptive transfer of MPP type 2 cells was sufficient to induce type 2 cytokine production and to confer protection in response to helminth infection (13). IL-25, together with IL-33, also induces expansion of nuocytes, primary source of IL-13 during helminth infection (14). Another epithelial cell-derived cytokine, thymic stromal lymphopoietin (TSLP) activates DC to drive Th2 immune responses (15). TSLP was recently reported to induce Th2 immunity by augmenting basophili production independently of IL-3 (16). Therefore, pathways toward Th2 immunity in vivo appear to be manifold.
We previously demonstrated that basophils are rapidly (and transiently) mobilized into the draining LN following Nippostrongylus brasiliensis (Nb) infection and that this recruitment requires IL-3 or IL-3Rβ (8). Yet, Nb specific Th2 immunity was efficiently induced in these mice, suggesting a basophil-independent Th2 response (8). Consistent with this, Th2 immune responses induced by primary Nb infection, papain immunization, passive immunoglobulin-mediated systemic anaphylaxis, and acute allergic lung inflammation were successfully induced without basophils, although basophils were found to be essential for protective memory responses against secondary Nb or tick infection, and chronic allergic skin inflammation (17-19). Therefore, mechanisms involved in Th2 immunity and basophils may be dependent on model systems used.
In this study, we report that papain-mediated basophil recruitment, similar to that mediated by Nb infection, is also IL-3-dependent and that the papain-induced Ag specific Th2 immune responses are not affected by the lack of IL-3 or IL-3Rβ, suggesting that basophils are dispensable for this process. Immunizing mice with nonviable parasites also induced IL-3-dependent basophil LN recruitment. Interestingly, Th2 responses induced by dead parasites were substantially greater than those induced by papain. Basophils recruited into the draining LN expressed little or no MHC II molecules compared to CD11c+ DC, and did not uptake Ag. In agreement with a recent report (7), FcεRI+ CD11c+ DC efficiently did uptake Ag. Furthermore, inducible basophil ablation did not affect Th2 immune responses. Unexpectedly, IL-4 produced by non-T cells was required for Th2 CD4 T cell responses to develop, whereas other innate type Th2 cytokines including IL-25 and TSLP were dispensable for the Th2 immune responses. Taken together, these results suggest a complexity of in vivo Th2 immunity that can arise without basophils or other innate type 2 cytokines.
Go to : 
METHODS
Mice
BALB/c wild type (WT), BALB/c IL-3-/-, and BALB/c DO11.10 TCR Tg mice were purchased from the Jackson Laboratory (Bar Harbor, ME). IL-3-/- mice and IL-3Rβ-/- mice, deficient in both βc and βIL-3 and G4 knock-in mice expressing GFP under the IL-4 promoter were previously described (20-22). IL-25R (IL-17RB)-/- mice were provided from Dr. Wenjun Ouyang (Genentech). Mcpt8DTR mice were previously reported (18). All experimental procedures were conducted according to the guidelines of the Institutional Animal Care and Use Committee.
Immunization
Mice were injected into the ear pinna with 100 µg chicken ovalbumin protein (Sigma, St. Louis, MO) plus 50 µg papain (Sigma) or 500 dead Nb parasites in a 10 µl volume. Seven days later, draining cervical LN was harvested and ex vivo restimulated with OVA323-339 peptide for 24 hours. IL-4 production was measured by ELISA or ELISPOT assay as described below. In some experiments, 250 µg anti-TSLP mAb (R and D system, Minneapolis, MN) was injected at days 0 and 3 post injection. For basophil depletion, mcpt8DTR or control mice were intravenously injected with diphtheria toxin (750 ng/20 g body weight; Sigma) prior to immunization.
Flow cytometry
Draining cervical LN cells and blood were examined for basophils. In brief, cells were stained with anti-FcγR (clone 93) and anti-CD45 (30-F11). Cells were ex vivo stimulated with PMA (CalBiochem, San Diego, CA) and Ionomycin (CalBiochem) for 4 hours. Monensin was added in the culture for the last 2 hours of stimulation. Cells were stained for intracellular IL-4 expression. Samples were acquired using a FACSCalibur or a LSRII (Becton Dickinson, Franklin Lakes, NJ) cytometer and analyzed using FlowJo (Treestar, Ashland, OR). All antibodies used were purchased from eBioscience (San Diego, CA).
ELISPOT and ELISA analysis
To measure T cell cytokine production harvested cells were stimulated with 10 µg OVA peptide for 24 hours. IL-4 production in the culture supernatant was measured by ELISA. In some experiment, IL-4 producing cells were determined by IL-4 ELISPOT assay. Abs used for both assays were purified anti-IL-4 (11B11) and biotinylated anti-IL-4 (BVD6-24G2). These Abs were purchased from eBioscience.
Data analysis
Statistical significance was determined by the Student's t-test using the Prism software (GraphPad Software, La Jolla, CA). p<0.05 was considered to indicate a significant difference.
Go to :
RESULTS
Papain immunization recruits circulating basophils to the draining LN via IL-3
In response to Th2-inducing allergen challenges or to parasite infections, circulating basophils are transiently recruited into the draining LN, where they are believed to play a key role in supporting the development of antigen specific Th2 type CD4 T cell responses (2). We previously reported that LN recruitment of circulating basophils after helminth infection was completely abolished in IL-3- or IL-3Rβ-deficient mice, indicating an IL-3-dependent process (8). First, we examined whether IL-3 also mediates basophil recruitment after immunization of papain, a prototype allergen shown to recruit basophils to the draining LN and induce an antigen specific Th2 immunity when coimmunized with the antigen (23). Groups of WT and IL-3-deficient mice were immunized with OVA plus papain and basophil recruitment in the draining LN was subsequently examined. Consistent with a previous report (23), injection of OVA protein plus papain induced a transient recruitment of FcγRhigh CD45intermediate basophils into the draining cervical LN of WT mice (Fig. 1). By contrast, such recruitment was completely absent in IL-3-deficient mice (Fig. 1). Therefore, papain-induced basophil LN recruitment also requires IL-3.
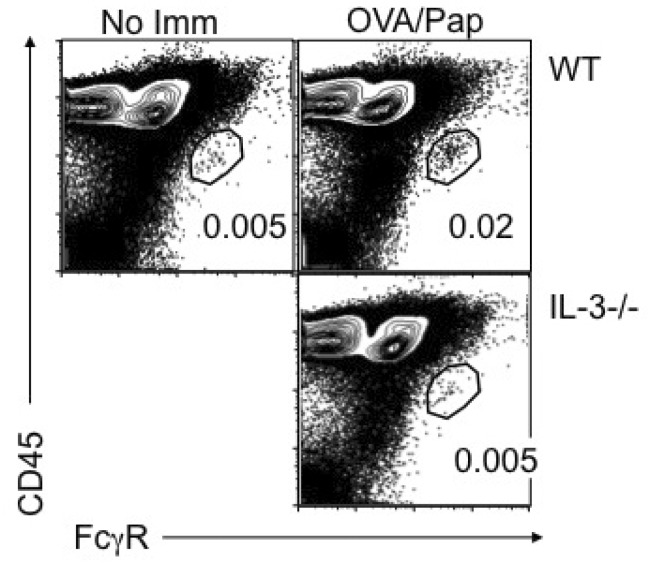 | Figure 1Papain immunization-mediated basophil LN recruitment requires IL-3. Groups of BALB/c and IL-3-/- mice were immunized with 100 µg OVA protein plus 50 µg papain. Draining cervical LN was collected 4 days post immunization. LN cells were examined for the existence of FcεRhigh CD45intermediate basophils. WT mice without immunization were included as negative controls. Data shown are representative of three repeated experiments.
|
IL-3 plays no role in papain-induced Th2 immune responses
Helminth-induced Th2 immunity normally develops in the absence of IL-3 or IL-3Rβ (i.e., in the absence of basophil recruitment into the draining LN) (8). In case of papain-induced Th2 immunity, however, the question of whether basophils are critical for the resulting Th2 immune responses remains unresolved. Since basophil LN recruitment was found to be absent in IL-3-deficient mice immunized with papain, we therefore examined OVA specific CD4 T cell responses in IL-3- or IL-3Rβ-deficient mice after immunization with OVA protein plus papain. First, OVA-specific DO11.10 TCR Tg CD4 T cells that are WT, IL-3-, or IL-3Rβ-deficient were adoptively transferred into WT recipients, and the recipients were subsequently immunized with OVA plus papain. As shown in Fig. 2A, comparable numbers of KJ1.26+ CD4 T cells were found in the draining LN following immunization regardless of the phenotypes of CD4 T cells. When proliferation was examined by CFSE dilution, the pattern of CFSE dilution was indistinguishable between WT, IL-3-/-, and IL-3Rβ-/- DO11.10 CD4 T cells (Fig. 2B). Therefore, the production of IL-3 or IL-3 responsiveness of CD4 T cells plays little or no role in Ag-induced T cell proliferation/expansion in vivo.
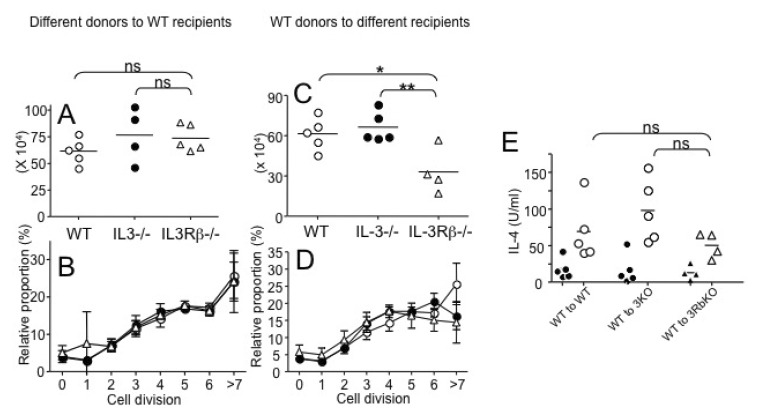 | Figure 2Roles of IL-3 and IL-3R during papain-induced Th2 immunity. (A, B) CD4 T cells were isolated from WT, IL-3-/-, and IL-3Rβ-/- DO11.10 mice, labeled with CFSE, and transferred into BALB/c recipients (3×106 cells per recipients). The recipients were then immunized with 100 µg OVA protein plus 50 µg papain into the ear pinnae. (A) Seven days post immunization, draining cervical LN cells were harvested. Total KJ1.26+ CD4 T cells were enumerated by FACS analysis. (B) CFSE dilution of KJ1.26+ CD4 T cells was also examined. (C~E) CD4 T cells were isolated from WT DO11.10 mice, labeled with CFSE, and transferred into WT, IL-3-/-, and IL-3Rβ-/- recipients (3×106 cells per recipients). (C) Total KL1.26+ CD4 T cells were enumerated as described above. (D) CFSE dilution was determined as described above. (E) Cervical LN cells were restimulated with OVA323-339 peptide for 24 hours and IL-4 production was determined by ELISA. Filled symbols are without peptide stimulation. Each symbol represents individually tested mouse. Similar results were obtained from two independent experiments. *p<0.05; **p<0.05;0.01.
|
Since recipient cells are fully capable of producing IL-3 in the experiments described above, we next transferred WT DO11.10 CD4 T cells into different types of recipients (WT, IL-3-/-, and IL-3Rβ-/-) to test whether IL-3 production or IL-3Rβ expression by the recipient cells plays any role in this process. As shown in Fig. 2C, DO11.10 CD4 T cells efficiently expanded in WT and IL-3-/- recipients. Interestingly, in IL-3Rβ-/- recipients the expansion was significantly reduced, which may be attributed to the defects in the expansion and maturation of antigen presenting cells in IL-3Rβ-/- mice (20). Nevertheless, CFSE dilution profiles of Ag specific T cells in all tested recipients was not significantly different (Fig. 2D), indicating that IL-3 production or IL-3Rβ expression of the endogenous recipient cells does not play a role in activating T cells, although IL-3Rβ expression of the recipients may contribute to the expansion of Ag specific T cells.
We then examined whether the inactivation of IL-3 or IL-3Rβ on the recipient derived cells including basophils affects papain-induced Th2 immune responses. Draining LN cells from immunized recipients were restimulated ex vivo with OVA peptide and IL-4 secretion was measured by ELISA. As shown in Fig. 2E, OVA-specific IL-4 secretion was similar regardless of the IL-3 (or IL-3Rβ) deficiency in the recipient cells. Likewise, IL-3-/- and IL-3Rβ-/- DO11.10 CD4 T cells transferred into WT recipients and immunized with OVA plus papain also secreted similar levels of IL-4 after OVA restimulation (data not shown). Therefore, basophils (or IL-3) seem to be dispensable in papain-induced Th2 immunity.
IL-3-independent Th2 immune responses after nonviable parasite Ag injection
We recently reported that injection of nonviable Nb larvae similarly induces basophil LN recruitment and robust Th2 responses and that basophil depletion using anti-FcεRI (MAR-1) mAb did not alter the responses (24). Similar to papain immunization or live helminth infection, dead Nb injection failed to recruit basophils into the draining LN in IL-3-/- mice (Fig. 3A). Interestingly, the magnitude of basophil recruitment by dead parasites was substantially greater than that induced by papain (Fig. 1 and 3A). Dead parasite immunization also induced stronger CD4 T cell IL-4 responses when compared to that induced by OVA/papain immunization (Fig. 3B and 3C). No IL-4 production was observed without restimulation or with irrelevant peptide stimulation (data not shown). Likewise, the lack of IL-3 from either transferred DO11.10 CD4 T cells and/or endogenous cells did not affect the generation of OVA-specific IL-4-producing T cells (data not shown). Therefore, these results suggest that in vivo Th2 immunity can arise without IL-3 (i.e., basophils).
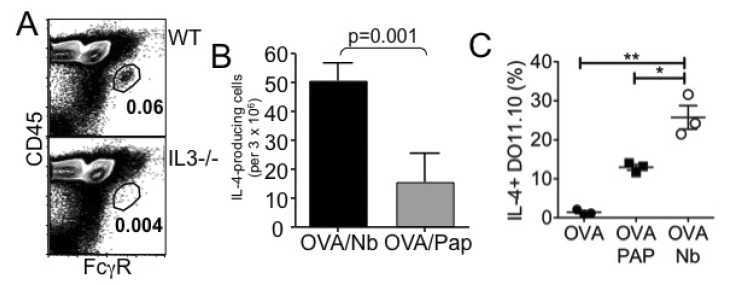 | Figure 3Nonviable N. brasiliensis immunization induces Th2 immunity. (A) WT or IL-3-/- mice were immunized with 100 µg OVA protein plus 500 dead L3 Nb parasites into the ear pinnae. Four days post injection, cervical draining LN was harvested and basophil recruitment was examined by FACS analysis. (B) Seven days post injection, cervical LN cells were harvested and in vitro stimulated with OVA peptide for 24 hours. IL-4-producing cells were determined by ELISPOT analysis as described in Materials and Methods. (C) Draining LN cells were in vitro stimulated with OVA peptide for 48 hours. Cells were stained for KJ1.26, CD4, and IL-4. DO11.10 CD4 T cell expression of intracellular IL-4 was determined by intracellular cytokine staining as described in Materials and Methods. *p<0.05; **p<0.01.
|
Basophil depletion did not affect Th2 immune responses
To ensure the dispensable role for basophils we utilized Mcpt8DTR mice in which basophils can be selectively depleted by DT administration (18). WT and Mcpt8DTR mice were transferred with DO11.10 CD4 T cells, injected with DT, and then immunized with OVA plus dead Nb. Basophil depletion was achieved by DT injection as measured by blood basophils (Fig. 4A). Consistent with earlier findings, basophil depletion by DT did not affect the development of IL-4-producing Ag specific CD4 T cell responses (Fig. 4B). Therefore, basophils are not critical for in vivo Th2 immunity.
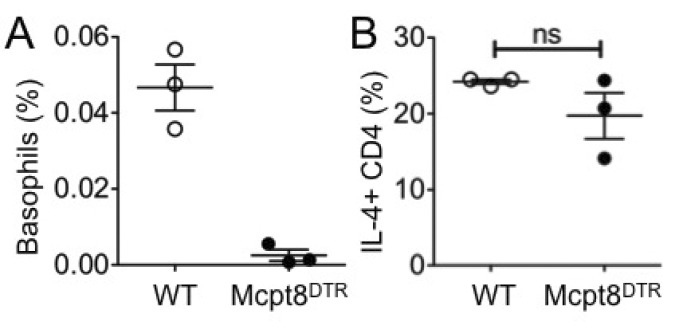 | Figure 4Th2 immune responses after basophil depletion. DO11.10 CD4 T cells were transferred into either WT or Mcpt8DTR mice. The recipients were injected with DT and immunized with OVA plus dead Nb. (A) Circulating basophils in the blood of the recipients were examined 7 days post immunization. (B) OVA specific IL-4-production was determined using draining LN cells harvested 7 days post immunization followed by in vitro 48 hours stimulation with OVA peptide. The proportion of IL-4 producing DO11.10 T cells was determined by FACS analysis. Each symbol represents individually tested mouse. WT, wild type; ns, not significant.
|
Basophils are not major antigen presenting cells in the draining LN
Whether basophils function as professional antigen presenting cells in the draining LN to induce Th2 immunity has been the matter of debate. Since a stronger Th2 immunity develops after immunization of dead Nb than of papain, we exploited this model system to compare cell types responsible for antigen presentation. Mice were immunized with OVA/papain or OVA/Nb, and MHC II expression of recruited basophils (CD45intermediate FcγRhigh, population II) and conventional antigen presenting cells (CD45high FcγRhigh, population I) was compared. As shown in Fig. 5A, MHC II expression of FcγR bearing antigen presenting cells (I) was very high; >90% of them were MHC IIbright. Moreover, no obvious difference was noticed between papain or dead Nb immunization. By contrast, MHC IIbright population was dramatically lower (~10%) in basophils (Fig. 5A) and most of the basophils did not express MHC II, similar to negative control CD45neg LN cells (population III). Therefore, basophils are not the major MHC II-expressing cells in the draining LN.
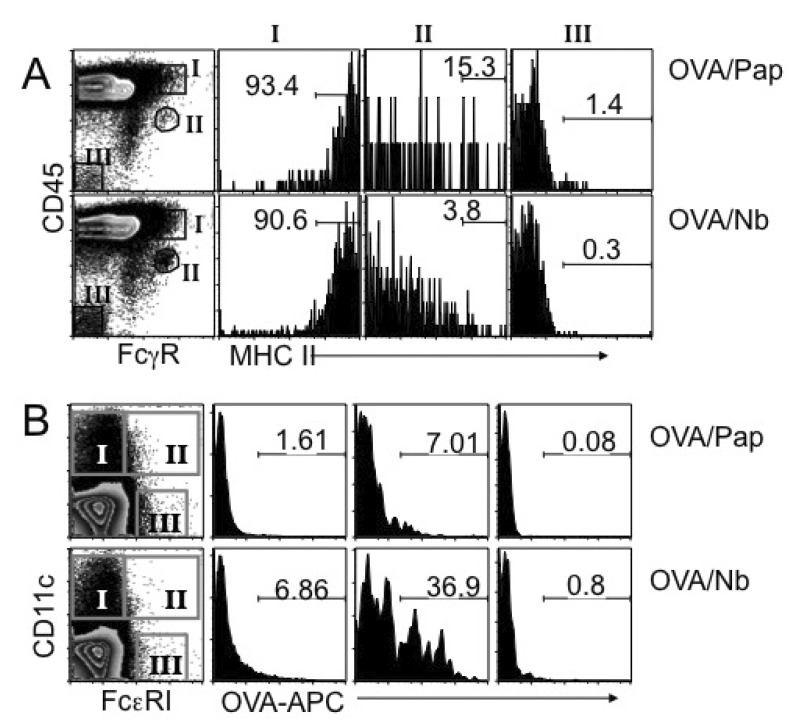 | Figure 5MHC II expression and Ag uptake of LN cells. (A) BALB/c mice were immunized with OVA plus papain or OVA plus dead Nb as indicated. Four days post immunization, draining LN cells were harvested, and stained for FcγR and CD45. MHC II expression on basophils (population II), FcγR/CD45high cells (population I) and non-lymphoid parenchymal cells (population III) were then examined. Shown are the proportions of MHC IIhigh cells among the indicated populations. (B) BALB/c mice were immunized with allophycocyanin (APC)-conjugated OVA protein plus papain or dead Nb as described above. Four days later draining LN cells were harvested, and stained for CD11c and FcεRI. Ag uptake of conventional CD11c+ DC (population I), FcεRI/CD11c+ inflammatory DC (population II), and FcεRI+ CD11c- basophils (population III) was examined by FACS analysis. The experiments were repeated twice with similar results.
|
It was demonstrated that endocytosis of soluble proteins by basophils is crucial for antigen presentation (3). In fact, ~15% of basophils are MHC IIbright in OVA/papain immunized mice, thus they may still be involved in antigen presentation to naïve T cells and in Th2 immunity. Alternatively, FcεRI-expressing 'inflammatory DC' subsets may function as key Th2-inducing antigen presenting cells (7,9). To address this possibility, mice were immunized with allophycocyanin (APC)-conjugated OVA plus papain or dead Nb. Fluorescent APC uptake of CD11c+ (I), FcεRI+ CD11c+ (II), and FcεRI+ CD11c- (III) cells was next analyzed. We found that APC-OVA uptake was predominantly found in FcεRI+ CD11c+ cells (Fig. 5B). By contrast, FcεRI+ CD11c- cells (III), which primarily include basophils, were not OVA-APC+ (Fig. 5B), strongly indicating that basophils are unable to uptake injected antigens. Importantly, OVA-APC uptake by FcεRI+ CD11c+ cells was dramatically enhanced when dead Nb was used instead of papain (Fig. 5B), which may result in a stronger Th2 immunity (Fig. 3B and 3C). OVA-APC uptake of conventional CD11c+ DC was substantially lower, although it was higher upon dead Nb injection (from 1.6% to 6.9%). Overall, these results provide strong evidence that dead Nb- and possibly papain-induced Th2 immunity is likely to be induced by antigen presentation of 'inflammatory DC' subsets that express FcεRI rather than basophils.
IL-4 but not IL-25 or TSLP is essential to generate dead Nb-induced Th2 immunity
To further investigate a mechanism(s) of how IL-3- and basophil-independent Th2 immunity develops we first tested the contribution of IL-4. WT DO11.10 CD4 T cells were transferred into WT and IL-4-deficient G4/G4 recipients. The recipients were then immunized with OVA plus dead Nb, and examined the development of OVA specific Th2 immune responses. Surprisingly, IL-4 production of OVA specific CD4 T cells was completely abolished in IL-4-deficient recipients, while robust Th2 immune responses were observed in WT recipients (Fig. 6A). Given that naïve CD4 T cells are capable of producing IL-4 sufficient for Th2 differentiation (25), these results strongly suggest that IL-4 produced by recipient non-T cells but not by Ag specific CD4 T cells in response to Nb/OVA immunization is required for the development of Th2 immune responses. Recently, innate type 2 cells such as nuocytes, MPPtype2, and NBNT cells have been implicated to play crucial roles in developing in vivo Th2 type immune responses particularly in the gut associated tissues (13,14,26). IL-25 and TSLP are candidate cytokines that activate those innate type 2 cells to promote Th2 immunity. In order to examine the contribution of innate type 2 immune components, we first used mice deficient in IL-25R (IL-17RB)-/- as recipients of DO11.10 CD4 T cells. The recipients were immunized with OVA plus dead Nb, and subsequent OVA-specific IL-4 responses were determined. As shown in Fig. 6B, the lack of IL-25R by recipient cells played no roles in generating IL-4-producing cells, indicating that IL-25-responsive innate Th2 cells may not be involved in this process. In addition, a potential role of TSLP was examined by treating WT recipients of DO11.10 CD4 T cells with neutralizing anti-TSLP mAb after immunization of OVA/Nb as previously reported (23). TSLP neutralization did not affect the generation of OVA-specific IL-4 responses (Fig. 6C). Therefore, neither IL-25 nor TSLP plays a role in the generation of IL-4-producing CD4 T cells in vivo and that IL-4 produced by recipient derived cells plays an essential role in promoting antigen specific Th2 immunity.
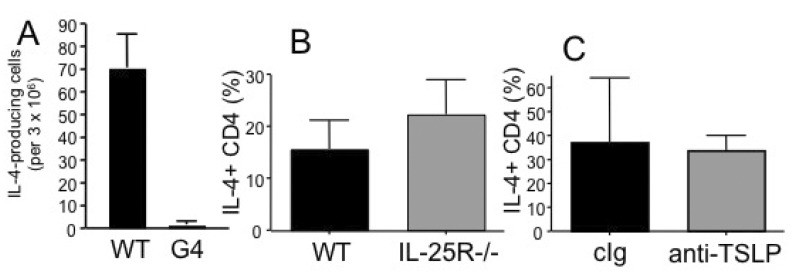 | Figure 6Roles of cytokine in dead Nb induced Th2 immunity. DO11.10 CD4 T cells were transferred into WT or IL-4-/- G4/G4 (A) or IL-25R-deficient (B) recipients. The recipients were immunized with OVA plus dead Nb as described above and IL-4-producing CD4 T cells were enumerated by ELISPOT assay as described above. (C) WT recipients of DO11.10 CD4 T cells were injected with neutralizing anti-TSLP mAb at days 0 and 4 of immunization. IL-4 production was examined as described above. Shown are the mean±STD of 3~4 individually tested recipients.
|
Go to :
DISCUSSION
Earlier findings that basophils are the major IL-4-producing innate cells have proposed an attractive hypothesis that they are the initiators of type 2 immune responses in vivo (23), which was further supported by several reports showing that basophils can function as both professional Ag presenting cells and the potent source of IL-4 (3-5). The model Ags used in these studies include: papain, the protease activity of which was critical for the Th2 induction (3), intestinal parasite Trichuris muris (27), and IgE-Ag complexes (28). This hypothesis has been challenged by several recent studies that provided compelling evidence that DCs rather than basophils play an indispensable role for the induction of Th2 immune responses in the model of Schistosoma mansonii infection and house dust mite Ag immunization (7,9). It was thus concluded that basophils are generally dispensable for the development of Th2 immunity in vivo, although they may still be the key effector cells mediating protection against certain pathogen infection (6,19).
Consistent with recent reports, the current study demonstrates that basophils are not necessary for papain-mediated Th2 immune responses as IL-4-producing CD4 T cells equivalently develop in the absence of IL-3 or IL-3Rβ, without which basophil LN recruitment is completely abolished (8). These findings strongly suggest that the entry of circulating basophils into the draining LN is not a requirement for the subsequent Th2 immunity to develop. Furthermore, relatively low expression of MHC II molecules as well as poor Ag uptake by basophils compared to those by FcεRI+ DCs further support the possibility that basophils are not involved in processing Ag and presenting it to naïve CD4 T cells. Likewise, inducible ablation of basophils did not affect the generation of IL-4-producing Ag specific CD4 T cells. Because very few basophils might still be found in the draining LN of IL-3- or IL-3Rβ-deficient recipients or even after basophil depletion, one might still argue that those few cells may be sufficient to function as Th2 inducers by presenting Ag and/or secreting IL-4. However, a recent study using IL-4:eGFP-KN2 transgenic mice in which IL-4 secretion can be traced by transgenic CD2 expression found that LN recruited basophils do not secrete IL-4 (6).
It is interesting to note that dead parasite-induced Th2 immune responses are completely abolished in IL-4-deficient recipients, because activated naïve CD4 T cells can produce IL-4 that can induce Th2 differentiation in an autocrine manner (25). However, our results clearly demonstrate that T cells are not the source of IL-4 at least in this system. Other innate type 2 cytokines such as TSLP and IL-25 can promote the development of Th2 immune responses particularly in the mucosal tissues through the induction of IL-4-producing innate type 2 cells. For example, IL-25 has been shown to induce IL-4 production in NKT cells, non-B/non-T (NBNT) c-kitpos cells, and MPPtype2 cells (11,12,29). However, we were unable to find Linneg c-kitpos IL-4 (GFP)-expressing innate type 2 cells within the draining LN (data not shown). Consistent with this, IL-4-producing CD4 T cell responses were efficiently generated in immunized IL-25R-deficient mice, suggesting that IL-25-responsive innate cells are not involved in this process. TSLP can promote Th2 immune responses (30); however, TSLP neutralization did not affect the generation of IL-4-producing CD4 T cells. IL-33 is an IL-1-related cytokine that is implicated in Th2-associated allergic inflammation (31). IL-33 is another cytokine that can stimulate various target cells including Th2 cells, mast cells, basophils, natural helper cells, and NKT cells (10). IL-33 primarily induces IL-5 and IL-13 expression (31). In the mediastinal LN of IL-33R-deficient mice induced for lung inflammation, CD4 T cell IL-4 expression was found reduced (32). It will be interesting to find if IL-33 is selectively expressed in the draining LN following papain or dead Nb immunization. However, dead Nb-induced Th2 immune responses were found unaltered in MyD88-/- recipients (data not shown), suggesting that IL-33 may not be involved in this process.
DC depletion severely disrupts the induction of Th2 immunity after Schistosoma egg Ag immunization or live Schistosoma mansonii infection (9). Consistent with this finding, in the model of house dust mite allergen challenge, basophil depletion only partially reduced allergen-induced Th2 immunity, while a novel DC subset that expresses the FcεRI was found to be essential for the induction of Th2 responses (7). Because IL-4 treated DC were shown to induce IL-4 production in DCs (33), it is possible that DC may produce small amount of IL-4 that acts on activated CD4 T cells to trigger Th2 differentiation. However, how DCs are induced to express IL-4 remains to be determined. Dead Nb associated molecules may induce IL-4 production in DCs. In conclusion, elucidating Th2 immunity generated through a basophil-independent pathway that requires IL-4 derived from non-T cells will add an additional complexity involved in the generation of type 2 immunity in vivo.
Go to :
 XML Download
XML Download