

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
NKT cells express both NK cell marker and T cell marker (1). NKT cells are distinguished type I NKT cell and type II NKT cells according to T cell receptor (TCR) usage. Type I NKT cells are called invariant NKT (iNKT) cells because they express only restricted TCR α chain (Vα14) and TCR β chain (Vβ2, 7 and 8), whereas type II NKT cells express various TCR αβ chains (2,3). NKT cells can help activation of other lymphocytes such as, T, B, NK and dendritc cells (3-6). It has been known that dedritic cells (DCs) play a pivotal role in antigen presentation to NKT cells (7). In contrast, resting B cells are known poor immunogenic antigen presenting cells (APCs) to NKT cells (8). However, B cell-NKT cell interaction can help reciprocally. NKT cells provide direct and indirect help for lipid and protein-specific B cells, respectively (9,10). Direct help from iNKT cells for lipid-specific B cells is dependent on IL-21 secreted from iNKT cells (11). High-density α-GalCer loaded B cells stimulate iNKT cells although the efficiency of stimulation is not comparable to dendritic cells (12). α-GalCer/peptide loaded B cells can induce peptide-specific cytotoxic CD8+ T cells and induced cytotoxic CD8+ T cells eliminate various kinds of tumor (12,13). In micro-environment of foreign antigen stimuli, DC and B cells cooperate to activate antigen-specific cytotoxic CD8+ T cells (14). Nevertheless, it remains to be resolved unknown NKT cells response upon co-operative stimulation of B cells and DCs.
In this study. we found that B cells induced Th1-skewed response of NKT cells in the presence of α-GalCer loaded DCs, regardless of exogenous antigen loading to B cells. We also demonstrated that regulation of NKT cell response by B cells stimulation was dependent on CD1d expression of B cells.
MATERIALS AND METHODS
Mice and reagents
Six- to eight-week-old C57BL/6 (B6), CD1d+/- and CD1d-/- mice (B6 background) were bred in facilities at Korea University. Mice were raised in a specific pathogen-free environment. The experimental protocols of this study were approved by the Institutional Animal Care and Use Committee of Korea University. Monoclonal antibodies specific for mouse TCRβ (H57), IFN-γ (XMG-1), IL-4 (11B11) and NK1.1 (PK136) were purchased from BD Biosciences. α-galactosylceramide (α-GalCer) were provided by Dr. Sanghee Kim (Seoul national univeristy).
DC, B and NKT cell isolation
Splenocytes was prepared as described previously (4). B cells were isolated from splenocytes via MACS by using anti-B220-conjugated magnetic beads (Miltenyi Biotec) after DC had been depleted by using anti-CD11c microbeads. DC were purified from splenocytes via MACS by using anti-CD11c microbeads. NKT cells were enriched from splenocytes by using CD4 microbeads.
In vitro activation of NKT cells
Purified B cells and DCs were loaded 10 ng/ml α-GalCer for 4 hr and they were washed extensively, then were co-cultured with NKT cells for 8 hr or 48 hr. After 8 hr or 48 hr, the amount of cytokines was measured by intracellular cytokine staining and ELISA.
Flow cytometry
Cells were washed and blocked with an anti-FcγRII/III mAb (2.4G2) for 15 min and then labeled for 30 min on ice with the appropriate mAbs. For intracellular cytokine staining, cells were fixed with a Cytofix/Cytoperm kit (BD Biosciences) according to the manufacturer's protocol. The stained cells were analyzed with a FACSCalibur flow cytometer using CellQuest software (BD Biosciences).
RESULTS AND DISCUSSION
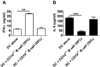
To test whether B cells can affect NKT cell stimulating activity of DCs, we co-cultured WT CD11c+ DC with NKT cells and B cells and then assessed the ability of B cells to activate NKT cells. ELISA analysis and intracellular staining demonstrated that IFN-γ from NKT cells was significantly increased, in contrast IL-4 was substantially reduced upon the addition of α-GalCer pulsed B cells compared to the response by DCs alone (Fig. 1). We next examined NKT response upon α-GalCer un-pulsed B cells addition. The co-culture of NKT cells with α-GalCer un-pulsed B cells also led to a significantly higher production of IFN-γ and lower production of IL-4 than that with α-GalCer pulsed DCs alone (Fig. 2A and 2B). Taken together, these results show that B cells promote Th1 response of NKT cells in the presence of antigen-loaded DCs, regardless of exogenous antigen pulse to B cells.
Although B cells promote Th1 response of NKT cells in the presence of antigen-loaded DCs, it was not clear whether NKT cells and B cells form directly cell-cell contact by CD1d-TCR interaction. To test whether CD1d expression on B cells was required to promote Th1 response of NKT cells in the presence of α-GalCer pulsed DCs, we isolated B cells from CD1d+/- or CD1d-/- mice and co-cultured them with DCs to activate NKT cells. Co-culture of NKT cells with CD1d+/- B cells led to a significantly higher production of IFN-γ and lower production of IL-4 than that with CD1d-/- B cells (Fig. 3). This data show that Th1-skewed response of NKT cells upon the addition of conventional B cells is dependent on CD1d expression of B cells.
Similarly, it had been reported that marginal zone (MZ) B cells amplify DC mediated-NKT cell activation (15). The cytokine patterns of NKT cells upon MZ B cells and DCs stimulation (15) was similar to our results, except IL-4 expression was also increased when α-GalCer loaded MZB cells was used. However, there was a contradicting report where conventional B cells could dampen iNKT cells activation in the presence of DCs (8). We are not sure at the moment what made these difference in NKT cell response but the density of B cells and concentration of α-GalCer might be causative factor since they used much higher number of B cells and higher concentration of α-GalCer to activate NKT cells compared to the experimental condition in this study. NKT cell-mediated cytokines milieu can be influenced by pattern of cellular composition of lymphocytes in specific inflammatory site. For example, NKT cells in tumor model promote Th1-skewed cytokine milieu, in contrast NKT cells in asthma model promote Th2-skewed cytokine milieu (4,6). In asthma model, the administration of α-GalCer ameliorated or worsened the disease symptoms by how to inject α-GalCer (16-18). A shift from Th2 to Th1-response by the administration of α-GalCer suppressed airway hyper-responsiveness (AHR) (17,18). In this study, we demonstrated that B cells promote Th1 response of NKT cells in the presence of antigen-loaded DCs. Thus, the administration of α-GalCer pulsed B cells and DCs may be a possible approach for the treatment of AHR. Eventually, the elucidation of optimal composition of APCs such as DCs and B cells to regulate finely the direction of NKT cell response will be the basis to apply NKT cell-based cell therapy.


 XML Download
XML Download