

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
MHC
major histocompatibility complex
APC
antigen presenting cell
CTLA-4
cytotoxic T-lymphocyte associated antigen 4
Fc
fragment crystallizable region
CD
cluster of differentiation
MLR
mixed lymphocyte reaction
ELISA
enzyme-linked immunosorbent assay
HRP
horseradish peroxidase
FITC
fluorescein isothiocyanate
PBS
phosphate-buffered saline
SDS-PAGE
sodium dodecyl sulfate polyacrylamide gel electrophoresis
TBST
tris-buffered saline tween-20
RT-PCR
reverse transcriptase-polymerase chain reaction
KDa
kilo-dalton
Kb
kilo-base
CPM
counts per minute
PVDF
polyvinylidene fluoride
INTRODUCTION
T cell costimulation is the most important concept in basic T cell immunology. Two types of signals provided by an antigen-presenting cell (APC) are essential for activation of T cells. Signal one is antigen-specific signal that is generated upon the engagement of T cell receptor (TCR) with the antigen (Ag) loaded in the major histocompatibility complex (MHC) on APCs. Signal two is produced by the binding between costimulatory receptors on T cells and their ligands expressed on APCs (1,2). Costimulation without TCR stimulation has no effect on T cells, but TCR stimulation with the absence of costimulation signal leads to T cell anergy (3).
One of the most important costimulatory signal enabling full activation of T cells is generated by binding between CD28 constitutively expressed on T cells and B7-1 or B7-2 on APC (4,5). In contrast, the interaction of B7 molecules with CTLA-4, a CD28 homologue provides inhibitory signals leading to down-regulation of the T cell response (6). CTLA-4 exhibits 10 to 100 times higher affinity for the B7 molecules than CD28 (7). To use as T-cell inhibitory agent, the extracellular domain of CTLA-4 was fused with Fc portion of IgG to generate the soluble form of CTLA-4 called CTLA-4Ig. In the transplantation field, CTLA-4Ig can prevent graft rejection including pancreatic islet, liver and cardiac transplantation (8-10). In addition, many approaches have been tried to improve functional efficacy of CTLA-4Ig through changing amino acid near binding site to B7 such as belatacept (11) or conjugating a carrier material to CTLA-4Ig such as liposomes (12).
In the case of conjugating a carrier molecule, antibody or enzymes were generally modified with chemical agents such as SATA (succinimidyl-s-acetylthioacetate) to crosslink with another maleimide-activated carrier material. In this study, we developed a modified form of CTLA-4Ig, termed as CTLA-4Ig-CTKC to facilitate direct conjugation without any chemical agents to maleimide-activated carrier molecule. In this report, we demonstrated that CTLA-4Ig-CTKC has same functional efficacy as CTLA-4Ig without any functional loss associated with single amino acid mutation.
MATERIALS AND METHODS
Animals
BALB/c and C57BL/6 mice were purchased from the Koatech (Kyunggi-do, Korea) and were kept at the specific pathogen-free animal facilities of the Seoul National University College of Medicine.
Cell cultures
A hybridoma cell line, 4F10 secreting anti-mouse CTLA4 antibodies (HB304, American Type Culture Collection, Manassas, VA, USA) and P815B7.1 cells, B7-1 transfected mouse mastcytoma cell line (TIB-64, American Type Culture Collection, Manassas, VA, USA) were cultured and maintained with high glucose Dulbecco's Modified Eagles Medium (DMEM, Gibco/BRL, Grand Island, NY, USA) containing 10% of heat-inactivated fetal bovine serum (FBS, Gibco/BRL, Grand Island, NY, USA) supplemented with 10 mM HEPES (Sigma-Aldrich Inc., St. Louis, MO, USA), 100 mM non-essential amino acids, 55µM β-mercaptoethanol (β-ME) and 50µg/ml gentamycin (Gibco/BRL, Grand Island, NY, USA).
CHO K-1 cells were also grown in DMEM for routine maintenance and in complete serum-free medium, Opti-CHO (Gibco/BRL, Grand Island, NY, USA) at mass culture for protein preparation. All of the cells were grown at 37℃ in humidified atmosphere with 7% CO2.
DNA construct and transfection
The DNA construct of mouse CTLA-4IgG3 was provided by Dr. Jeffery A. Bluestone. All plasmid DNA were prepared by using the DNA preparation kit (Genenmed, Seoul, Korea). Point mutation of IgG3 c-terminus was proceeded by PCR method to make c-terminal residue from AAA (lysine) to ACA (cysteine). PCR reaction was performed using Han-pfu polymerase (Genenmed, Seoul, Korea) with following two primers; sense, 5'-GTG GTA CCT TTA ATG AAA-3'(18 mer), antisense, 5'-GCT CTA GAG CTG TTC TCA ACA ACC AGG GGA GCG A-3'(34 mer) which contains codon for cysteine (bold character). This reaction was set up as follows: 1 ng of CTLA-4Ig DNA, 0.2µM of sense and anti-sense primer, 5µl of 10× reaction buffer (supplied with Han-pfu polymerase), 10 mM dNTP, 1µl Han-pfu polymerase, in a final volume of 50µl. Reaction cycle was as follows: 94℃ 3 min for pre-denaturation, 35 cycles of 94℃ for 1 min, 58℃ for 1 min, 72℃ for 1 min and 72℃ 10 min for post elongation. The PCR product was digested by Kpn I and XbaI and ligated to pcDNA3.1 (Invitrogen Life Technology, Carlsbad, CA, USA). DNA construct was then transformed into E. coli DH5 alpha followed by preparing plasmid DNA. Mutation site was confirmed by DNA sequencing analysis. CHO cells were collected at log-phase growth, and electroporation was performed as follows. Each 10µg of both pcDNA3.1-CTLA4 wild type and pcDNA3.1-CTLA-4IgG3-CTKC was mixed with each 5×106 cells in a 0.4-cm electroporation cuvette and electroporated at 25µF and infinite resistance using 300 volt in Gene pulser X cell system (Bio-RAD Laboratories, Hercules, California, USA) and plated into 100 mm culture dish.
In vitro transcription and translation
The TNT® Quick Coupled Transcription/Translation System (Promega, Madison, MI, USA) was used for in vitro transcription/translation and Transcend™ Non Radioactive Translation Detection System (Promega, Madison, MI, USA) was used for detecting the translation products. One µg of both pcDNA3.1-CTLA4Ig and pcDNA3.1-CTLA4IgCys was transcribed and translated in a 50µl reaction mixture containing 40µl of TNT®-T7 quick master mix, 1 mM methionine, and 2µl of Transcend™ Biotin-Lysyl-tRNA at 30℃ for 90 minutes. The products of reaction were separated by SDS-PAGE and transferred to a PVDF membrane. The transfer was performed in the transfer buffer containing 193 mM glycine, 25 mM Tris and 20% methanol for 2 hours at 100 V at 4℃. The membrane was then submerged in incubation buffer containing 5% non-fat dried milk and 0.1% Tween-20 in 20 mM Tris/HCl (pH 7.3)/137 mM NaCl (TBST) and left for 3 hours at room temperature. The detection antibody, streptavidin-HRP conjugate was diluted 1:10,000 in TBST and added to the membrane followed by incubation for 2 hours at room temperature. The development of positive signal was performed using Supersignal West Pico Chemiluminescent substrate (Pierce, Thermo scientific, Rockford, IL, USA).
Transfection, selection and mass culture
One day after incubation of electroporated CHO cells, cells were harvested and splitted into 20 plates of 100 mm dish with media containing 2 mg/ml of G418 (Geneticin, Gibco Laboratories, Grand Island, NY, USA) to select stable clones. For selecting stable clones, transfected cells were incubated for 2~3 weeks in the presence of G418 until cell colonies were visible. Ninety six cell colonies were picked on an inverted light microscope using micropipette and transferred to 96-well culture dish (Nunc, Rochester, NY, USA) with 0.2 ml of culture media containing 2 mg/ml of G418. Individual colony was cultured and a few clones were sorted out by high expression level of the secreted protein. The sorted cells were sub-cultured and adapted in complete serum free medium, Opti CHO (Gibco/BRL, Grand Island, NY, USA). The selected cells (3×107) were transferred to Triple flask (Nunc, Rochester, NY, USA) for mass culture with 250 ml of Opti CHO medium.
4F10 sepharose bead preparation
The hamster anti-mouse CTLA-4 mAb 4F10 was purified from hybridoma cell culture supernatants using standard protein A-sepharose chromatography. One gram of CNBr activated sepharose Fast flow (GE Healthcare, San Jose, CA, USA) was swollen with 1 mM HCl for 15 min at room temperature. The swollen gel was transferred to sintered glass funnel and washed with 200 ml of 1 mM HCl. After washing gel with 5 ml coupling buffer (0.1 M NaHCO3, 0.5 M NaCl), gel was transferred to tube containing 20 mg of anti-mouse CTLA-4 monoclonal antibody, 4F10, in coupling buffer and incubated for 2 hours at room temperature with constant rotation. After coupling reaction, gel was incubated with blocking buffer (0.2 M Glycine, pH 8.0) to block the excess active group for 2 hours at room temperature. This gel was then poured into sterile empty chromatography column followed by washing with the four times alternating between low pH wash buffer (0.1 M acetic acid, 0.5 M NaCl) and coupling buffer and stored at 4℃ with PBS (137 mM NaCl, 2.7 mM KCl, 8 mM Na2HPO4, 1.5 mM KH2PO4, pH 7.4) containing a preservative (sodium azide 0.05%) until use.
Measurements of quantification of secreted protein
To quantify the amount of CTLA-4Ig during selection and mass culture, sandwich-enzyme-linked immunosorbent assay (ELISA) was performed by following antibodies; Anti-CTLA4 monoclonal antibody (4F10) as capture antibody and HRP conjugated anti-mouse IgG3 antibody as detecting antibody.
Two µg/ml of anti-CTLA4 monoclonal antibody diluted in PBS was coated on 96-well MaxiSorp™ ELISA plate (Nunc, Rochester, NY, USA) and incubated overnight at 4℃ and washed 3 times with PBS containing 0.05% of Tween-20 (PBST) (Sigma-Aldrich Inc., St. Louis, MO, USA). After blocking of each well with 5% skim milk (Difco, Spark, MD, USA) in PBST for 3 hours at room temperature, diluted sample (1:100~1:100,000) was added in each well for 2 hours at room temperature and washed 5 times with PBST. The HRP conjugated anti mouse IgG3 (GeneTex, Irvine, CA, USA) diluted in 1:5,000 was added each well for 2 hours at room temperature and washed 5 times with PBST. One hundred µl of TMB substrate solution (eBioscience, San Diego, CA, USA) was added to each well for 5 minute and same volume of 0.18M H2SO4 was used for stopping color reaction. The plate was read at an absorbance of 450 nm.
Preparation of CTLA-4Ig
All of CTLA-4Igs in either wild or mutant form were produced in our laboratory. Before handing with mutant form of CTLA-4Ig, we have done many in vitro and in vivo experiments with wild type CTLA-4Ig. When purifying this wild type form, protein G affinity column was enough for obtaining pure CTLA-4Ig. However, mutating c-terminal lysine to cysteine, a significant amount of mutant form of CTLA-4Ig got through protein G column, indicating single amino acid mutation cause some structural change to yield lower binding affinity to protein G. Therefore, we prepared a 4F10 sepharose bead by ourselves and devised a new purification method. To rule out any structural and/or functional change associated with a new purification method, we have purified wild type and mutant form of CTLA-4Ig described in this manuscript.
Total culture supernatant was harvested from several Triple flask cultures and centrifuged at 12,000 g at 4℃ by the Avanti J-E high speed centrifuge (Beckman Coulters, Fullerton, CA, USA). Supernatant was carefully harvested and was then filtered by 0.22µm sterile top filter (Millipore, Billerica, MA, USA).
After 4F10-sepharose column was equilibrated with 3 bed volumes of PBS, the filtered sample was applied to the column and allowed to flow completely into the gel. After a few turn over of flow through column, 30 ml of Immunopure® IgG elution buffer (Pierce Biotech nology, Rockford, IL, USA) was added to column and collected 1.0 ml fractions in 1.5 ml tube which contains 100µl of 1M Tris, pH 8.0. The collected fractions were quantified by measuring the absorbance at 280 nm by the NanoDrop, ND-1000 (Thermo Scientific, Wilmington, DE, USA) and concentrated with the Amicon Ultra centrifugal filter (Millipore, Billerica, CA, USA). Concentrated fraction was quantified by BCA assay kit (Pierce Biotechnology, Rockford, IL, USA).
Western blotting
Purified CTLA-4Ig proteins (5~10µg) were loaded in SDS-PAGE under two different conditions, reducing and non-reducing condition and electro-transferred to PVDF membrane. Next, the PVDF membrane was incubated with 5% skim milk in PBST for 2 hours at room temperature and probed with 10µg of anti-mouse CTLA4 antibody, 4F10, in 15 ml of PBST containing 2% skim milk. After 2 hours at room temperature or overnight incubation at 4℃ with gentle shaking, 3 times of washing with PBST was followed by adding the HRP-conjugated anti Armenian hamster IgG (Santa Cruz Biotechnology, Delaware, CA, USA) as detection antibody (1:5,000 dilution) with same buffer for 2 hours at room temperature. The positive signals were developed with adding 2 ml of Enhanced Chemiluminescent (ECL) reagents (Pierce Biotech nology, Rockford, IL, USA).
Islet isolation and transplantation
Islet isolation was performed as described previously (13). Briefly, Balb/c mice were sacrificed, and the pancreas was injected with Hank's balanced salt solution containing 0.5 mg/ml of collagenase P (Roche, Mannheim, Germany) via the pancreatic duct until pancreas was inflated and then incubated in 37℃ for 20 minutes. Islets were purified on Euro-Ficoll gradients by centrifugation at 900 × g for 15 minutes and followed by incubation with complete RPMI medium (Invitrogen, Carlsbad, CA, USA) for overnight culture. Next day, islets were hand-picked and were used for the transplantation. Five hundred hand-picked islets were transplanted under the left kidney capsule of C57BL/6 mice rendered diabetic by streptozotocin (STZ). Following islet transplantation, 50µg of CTLA-4Ig or CTLA-4Ig-CTKC was injected via intra peritoneum every other day for 14 days.
RESULTS
Generation of DNA construct of both CTLA-4 IgG3 and mutant
Mouse CTLA-4Ig construct was kindly provided by Dr. Bluestone in UCSF, while mutant form of CTLA-IgG3 was made by point mutation with PCR method. Amino acid at the end of CH3 domain of IgG3 is lysine, but this residue was changed to cysteine by mutagenesis. According to construct diagram, nucleotide sequence at this site was changed from AAA (Lys) to TGT (Cys) (Fig. 1A). The size of PCR product containing mutation site was confirmed by 1% agarose gel electrophoresis as 1.2 Kb band in Fig. 1B. This construct was subcloned into the eukarytotic expression vector, pcDNA3.1, and full DNA sequence of PCR product was confirmed by DNA sequencing analysis. The protein expression of pcDNA3.1-CTLA-4IgG3 was further confirmed by the TNT® Quick Coupled Transcription/Translation System (Promega, Madison, MI, USA), SDS-PAGE and Western blotting. The results showed that both wild type and mutant type proteins yielded about 45 KDa protein band (Fig. 1C).
Purification of CTLA-4Ig and CTLA-4Ig-CTKC
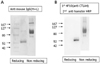
To make a stable cell lines, the purified DNA constructs were transfected into CHO cell lines with electroporation method. Several clones exhibiting high levels of secretion of each CTLA-4Ig or CTLA-4Ig-CTKC proteins were established. After establishment of stable cell lines, cells were adapted to serum-free CHO cell medium to start mass culture. Total 6 liters of culture supernatant were harvested in every purification cycle. The mutant form of CTLA-4Ig was purified by using anti-CTLA4 mAb, 4F10, conjugated sepharose bead because of its low binding affinity to protein G sepharose beads (data not shown). Purified protein was identified as 55 KDa under reducing condition and 110 KDa under non-reducing condition in SDS-PAGE (Fig. 2). These results indicate that the secreted proteins from cells have a divalent CTLA4 molecule and slight increase in the size of protein might be due to glycosylation of target protein in CHO cells.
B7-1 binding assay of purified CTLA-4Ig-CTKC
To test whether purified CTLA-4Ig-CTKC binds to its target molecules, B7-1 (CD80), varying amounts of purified protein were added to B7-1 expressing P815 cell line, P815B7-1. As shown in Fig. 3, CTLA-4IgG3-CTKC completely bound to B7.1 at 50 ng/ml of concentration. The mean fluorescence intensity of both CTLA-4Ig and CTLA-4Ig-CTKC increased in a dose-dependent manner from 50 ng/ml to 1µg/ml of proteins. Especially, CTLA-4Ig-CTKC had a similar binding efficacy compared with CTLA-4Ig although binding affinity of CTLA-4Ig-CTKC was slightly lower than that of CTLA-4Ig (Fig. 3 and 4). These results showed that CTLA-4Ig-CTKC could bind its ligand similarly as CTLA-4Ig.
The inhibitory effects of CTLA-4Ig-CTKC on MLR response
To investigate a possible effect of CTLA-4Ig in blocking the B7 molecule on APC, both CTLA-4Ig and CTLA-4Ig-CTKC were added to MHC-disparate allogeneic (BALB/c and C57BL/6) MLR condition. 3H-thymidine uptake by proliferating T cells from C57BL/6 mice splenocytes was measured. Count per minute (CPM) values increased over 50,000 at 72 hours in time-dependent manner in untreated control group (0µg/ml). However, when CTLA-4Ig or CTLA-4Ig-CTKC was treated (0.5~6.0µg/ml), proliferative responses were dramatically decreased in a dose-dependent manner (Fig. 4). This result demonstrated that CTLA-4Ig-CTKC also had inhibitory function on T cell proliferation in vitro similarly as CTLA-4Ig.
The CTLA-4Ig-CTKC was able to prolong islet graft survival similarly as CTLA-4Ig
To further confirm about the functional efficacy of CTLA-4Ig-CTKC, both CTLA-4Ig and CTLA-4Ig-CTKC were used in the allogeneic pancreatic islet transplantation model. Diabetic C57BL/6 mice received the isolated islet from Balb/c mice. As shown in Fig. 5, the animal treated with CTLA-4Ig-CTKC maintained blood glucose level over 100 days similarly as CTLA-4Ig-treated group. This result showed that CTLA-4Ig-CTKC can effectively inhibit CD28-mediated costimulatory signal in alloantigen specific T cells in vivo.
DISCUSSION
In this study, CTLA-4Ig-CTKC was generated by point mutation at the end of Fc portion in IgG3 and its immunological function was examined in vitro and in vivo. Our results showed that single amino acid substitution in CTLA-4Ig-CTKC does not affect its function as revealed by similar affinity to B7-1 molecule, prevention of T cell activation in allogeneic MLR reaction and prolongation of allogeneic islet graft in transplantation model.
CTLA-4Ig has been developed as an effective CD28 antagonist. The mechanistic basis for CTLA-4Ig function in vivo has increasingly become clear. CTLA-4Ig has been shown to be effective in the treatment of autoimmune diseases and thus approved by the Food and Drug Adminstration (FDA) (Abatacept, Bristol-Myers Squibb, NY, USA) for the treatment of rheumatoid arthritis (14-16). More recently, an improved version of Abatacept termed as LEA29Y with respect to higher affinity and avidity than CLTA-4Ig (Belatacept, Bristol-Myers Squibb, NY, USA) was developed and its efficacy was tested in the phase III clinical trial study in kidney transplantation (17-21). Another approach to increase affinity and avidity of CTLA-4Ig reported so far was to conjugate CTLA-4Ig on the surface of stealth liposomes such as PEG-PE liposomes (12). In that study, CTLA-4Ig conjugated to liposomes had been shown to improve the avidity of CTLA-4Ig binding to B7 molecule by its multivalent effect, but most of the injected CTLA-4Ig liposome was trapped in liver and spleen via non-specific binding to Fc receptor (12). The usage of mouse IgG3 for CTLA-4Ig might be a solution to overcome this problem because Fc portion of mouse IgG3 was known to have a low affinity for Fc receptors (22). In addition, another way to improve CTLA-4Ig's efficacy would be to give unidirectional orientation on the liposomes. To this end, a mutant form of CTLA-4IgG3, termed as CTLA-4Ig-CTKC, was developed by changing the c-terminal end of IgG3 from lysine to cysteine. We suggest that this cysteine is directly reactive to maleimide-activated carrier materials without chemical modification of CTLA-4Ig such as succinimidyl-s-acetylthioacetate (SATA) for creating free thiol group (-SH).
In protein engineering, a single amino acid substitution may yield problems such as loss of binding affinity, irreversible aggregation, and instability. Therefore, mutant proteins should be tested whether they maintain binding affinity and function. CTLA-4Ig-CTKC actually had a lower binding affinity to protein G sepharose column compared with CTLA-4Ig, leading us to find another purification strategy. Fortunately, CTLA-4Ig-CTKC was successfully purified by using 4F10-sepharose beads made in house and its immunological function was examined in vitro and in vivo. CTLA-4Ig-CTKC was able to suppress proliferation of T cells in MLR similarly as CTLA-4Ig and also bound B7-1 molecule in the binding assay although binding affinity was slightly lower than that of wild type. This propensity was also observed in the allogeneic islet transplantation model. It is well known that CTLA-4Ig induces T cell-mediated tolerance and thus achieve long-term graft survival in the transplantation model. As shown in Fig. 5, CTLA-4Ig-CTKC-treated mice also maintained blood glucose level over 100 days. This result clearly demonstrated that the functional efficacy of CTLA-4Ig-CTKC was not altered and thus can be used for further application procedure.
In conclusion, a modified form of CTLA-4IgG3, CTLA-4Ig-CTKC, has a high affinity to B7-1 and also inhibits T cell proliferation. It is possible that CTLA-4Ig-CTKC can be directly conjugated to maleimide-activetaed carrier materials such as liposomes. Further study is needed to test whether conjugated CTLA-4Ig-CTKC liposomes would have more potent immune suppressive ability than CTLA-4Ig.




 XML Download
XML Download