

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
APC
antigen-presenting cell
C5aR
complement C5a receptor
ELISA
enzyme-linked immunosorbent assay
CLSM
confocal laser scanning microscope
ELISPOT
enzyme-linked immunospot
FAE
follicle-associated epithelium
FMDV
foot-and-mouth disease virus
LP
lamina propria
PP
Peyer's patch
SC
secreting cell
SE
standard error
SIgA
secretory IgA
SPL
splenocyte
UEA-1
Ulex europaeus agglutinin 1
WGA
wheat germ agglutinin
INTRODUCTION
Foot-and-mouth disease virus (FMDV), belonging to the genus Aphthovirus, is arguably the most dreaded virus of ungulates worldwide (1). Economic impact of FMDV is enormous because millions of animals are lost each year, and it imposes limitation on economic activities, including tourism and trade (2). Current inactivated FMDV vaccine requires the growth of large amounts of virulent FMDV and purification procedure for high concentrations of FMDV antigen is suggested as a causative factor in FMDV transmission when it is not properly controlled (3). FMDV-specific IgA production was reported to correlate with protection against viral challenge, which suggested the importance of mucosal IgA response in vaccination (4). Further, given that the cost of vaccination is a great burden in animal farming industry, eliciting mucosal IgA immune response by applying oral mucosal vaccine is an ideal strategy in developing effective vaccine against FMDV infection (3).
Needle-free oral vaccination is safe and easy to administer. In addition, it could induce antigen-specific immune response in both systemic and mucosal compartments (5). Considering that 90% of all pathogen infections occur in mucosal areas, it is conceivable that mucosal vaccination capable of establishing protective immunity in this frontline of pathogen infections could overcome limitations of current injection-based vaccines (6). Nevertheless, intrinsic characteristics of the mucosal immune system impose the great task to achieve efficient immune response induction through oral mucosal immunization. For example, mucosal epithelium covering the aerodigestive and urogenital tracts constitutes a special immune system that maintains mucosal homeostasis by restricting the influx of luminal antigens and dominantly inducing tolerance (7). Although these characteristics of mucosal environment can prohibit pathogen infections and immune response induction against unwanted luminal antigens, such as food antigens, at the same time, it hampers successful oral mucosal vaccination. In order to overcome obstacles to develop the effective oral mucosal vaccine, M cells are considered a good target to deliver vaccine materials and stimulate immune responses (8). M cells localized in follicle-associated epithelium (FAE) of Peyer's patch (PP) are specialized epithelial cells to take up luminal antigens through the thin glycocalyx layer and short/irregular microvilli, and to express specific receptors, such as GP2 and the complement C5a receptor (C5aR) (9-11). In addition, M cells are closely involved in the induction of antigen-specific immune response through several antigen-presenting cells (APCs) localized in an intracellular pocket which could come to contact with antigens transcytosed through M cells. APCs loaded with antigens stimulate T cells and induce the T cell-dependent secretory IgA (SIgA) production which determines the longevity of antigen-specific plasma cells and memory cells (12). Consequently, many studies on the development of effective oral mucosal vaccine have been concentrated on the development of an antigen delivery system to M cells.
Previously, we reported M cell-targeting ligand Co1 which was selected by phage display library screening against in vitro human M-like cells (9). Application of the ligand in a mouse oral mucosal immunization model confirmed the adjuvant effect of the ligand through effective targeting of conjugated antigen into M cells and induction of the Th2-skewed immune-stimulating effect (9). In this study, we applied the Co1 ligand to VP1 of FMDV which contains neutralizing epitopes and tested its efficacy in enhancing induction of VP1-specific immune response in both systemic and mucosal compartments after oral administration in mice. It was confirmed that Co1 ligand-conjugated VP1 of FMDV was targeted to M cells in ileal PPs. It was also verified that oral priming with VP1-Co1 effectively elicited Th2 cell-mediated VP1-specific IgA responses in PP and lamina propria (LP) after systemic challenge with VP1. It also induced T cell-mediated VP1-specific systemic immune response without inducing of systemic tolerance. Collectively, it is suggested that M cell-targeting ligand Co1 could be used as a mucosal adjuvant to enhance the delivery of VP1 antigen into mucosal immune inductive site and immune stimulation against the conjugated VP1 antigen in an oral mucosal vaccine model against FMDV infection.
MATERIALS AND METHODS
Experimental materials
All reagents used in this study were purchased from Sigma Chemical Co. (St. Louis, MO, USA) until specified otherwise. BALB/c mice were purchased from Charles River Technology through Orient Bio (Sungnam, Korea).
Production of recombinant antigens
To produce recombinant antigens either with or without Co1 ligand-conjugation, the VP1 gene was amplified from the pMYF18 plasmid vector containing the codon-optimized VP1 gene of FMDV which was kindly provided by Dr. M.-S. Yang (Chonbuk National University, Jeonju, Korea). The forward primer sequence used to amplify genes for FMDV VP1 alone and the Co1 ligand-conjugated VP1 was 5'-CCC AAA GCA TGC ACC ACC TCC ACA GGT GAG TCC-3', in which underlined letters represent the sequence for the SphI site. The reverse primer sequences used to amplify genes for FMDV VP1 alone and Co1-conjugated VP1 are 5'-GGG TTT GTC GAC CTA GAG GAG CTG CTT CAC AGG AGC-3' and 5'-GGG TTT GTC GAC CTA CGG CAG AGG CGA CCG CGC CGG CAG CTG ATG AAA CGA GAG GAG CTG CTT CAC AGG AGC-3', respectively, in which underlined and italic letters represent the sequences for the SalI site and Co1 ligand, respectively. Amplified gene products were digested with SphI and SalI, and cloned into the pQE30 expression vector (Qiagen, Hilden, Germany). Recombinant proteins were expressed using M15 expression hosts and purified as described in previous studies and as suggested by the manufacturer (9).
In vitro M cell-targeting assay
To monitor the interaction between antigens and M cells, PPs were obtained from the ileum and incubated with either VP1 or VP1-Co1 recombinant protein. After washing with ice-cold PBS, PPs were fixed by 4% paraformaldehyde, blocked with 5% BSA and 0.1% glycine in PBS, and then stained with rhodamine-conjugated Ulex europaeus agglutinin 1 (UEA-1, Vector Laboratories, Burlingame, CA, USA), eFlour 405-conjugated wheat germ agglutinin (WGA, Invitrogen, Carlsbad, CA, USA), and anti-VP1 antibody (kindly provided by Dr. K. Kunert, University of Pretoria, Pretoria, South African Republic), followed by FITC-conjugated secondary antibody. The specimens were then monitored by using a confocal laser scanning microscope (CLSM; LSM 510 META; Carl Zeiss, Thornwood, NY, USA).
Analysis of antigen-specific immune responses
A total of 100µg recombinant protein was orally introduced into 5 week-old female BALB/c mice, 5 mice per group, without anesthesia, once per week for 6 weeks. At 20 days after the last oral administration, mice were boosted by intraperitoneal injection of recombinant VP1 antigen without Co1 ligand conjugation. To measure the level of VP1-specific IgA, fecal extracts were collected at 5 days after the systemic challenge and analyzed by enzyme-linked immunosorbent assay (ELISA) as described previously (9). Lymphocytes were also prepared from the spleen, PP, and LP at 10 days after the boost immunization and subjected to measure VP1-specific IgG-, IgA- and indicated cytokine-secreting cells (SCs) through enzyme-linked immunospot (ELISPOT) assay, as described previously (9). Antigen-specific splenocytes (SPLs) proliferation was determined by the thymidine incorporation assay as described previously (9).
RESULTS
M cell-targeting ligand-conjugated VP1 interacted efficiently with M cells in FAE of PP
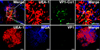
In oral mucosal immunization, introduction of antigen into the immune inductive site is prerequisite to induce efficient antigen-specific immune responses (13). Our previous study demonstrated that Co1 peptide (N-SFHQLPARSPLP-C) could be used to deliver conjugated model antigen into M cells in PP, a mucosal immune inductive tissue (9). To test whether this ligand could be applied in oral mucosal vaccine against FMDV infection, Co1 ligand was initially conjugated to C-terminus of VP1 antigen and its interaction with M cells was monitored as described in Materials and Methods (Fig. 1). M cells were detected as binding with UEA-1 (red), but not with WGA (blue), and VP1 proteins were identified by using anti-FMDV polyclonal antibody and then by using FITC-conjugated anti-rabbit IgG (green). As shown in Fig. 1, Co1 ligand-conjugated VP1 was localized in M cells of PP (yellow on the merged image in the upper panel), while VP1 protein without Co1 ligand was not detected in PP (lower panel). Thus, it was shown that M cell-targeting ligand Co1 conjugation enhanced the delivery of ligand-conjugated VP1 antigen into the mucosal immune inductive site.
Oral administration of ligand-conjugated antigen induced the enhanced mucosal immune response against the ligand-conjugated VP1 antigen
On the basis of the observed antigen-targeting ability of Co1 ligand into M cells of PP, we assumed that oral administration of Co1 ligand-conjugated VP1 may induce efficient mucosal and systemic immune responses against VP1. To confirm the possible practical application of M cell-targeting ligand Co1 into pathogenic antigen, we initially assessed the level of mucosal IgA after oral administration of the antigen because it has previously been known that mucosal IgA response is associated with protection of the host against FMDV infection (4). When we measured the level of VP1-specific IgA in fecal extracts which were collected after systemic boost immunization with VP1 alone after oral administration of 100 µg of either VP1-Co1 or VP1 alone once a week for 6 weeks, the level in mice fed Co1-conjugated VP1 was more than 4-fold higher than that in mice fed VP1 alone (Fig. 2A). This enhanced level of fecal IgA induced after oral administration of Co1-conjugated VP1 was supported by an increased number of VP1-specific IgA SCs in LP lymphocytes (Fig. 2B).
Induction of antigen-specific mucosal immune response which is closely associated with the T cell-dependent IgA production pathway occurs in germinal centers (12). Therefore, we tested whether mucosal IgA response enhanced by Co1 ligand-conjugation was associated with antigen-specific T cell responses against the antigen by characterizing the pattern of Th1/Th2-type cytokine expression in lymphocytes prepared from PPs through ELISPOT (Fig. 3). Oral administration of Co1-conjugated VP1 decreased the number of IFN-γ SCs about 8-fold and increased the number of IL-6 SCs about 3-fold in mucosal immune induction sites after in vitro VP1 stimulation (Fig. 3A). Oral administration of Co1-conjugated VP1 also increased the number of lymphocytes secreting Th2-type cytokines in LP, the mucosal effector site, more than 4-fold compared to that after oral immunization with VP1 alone (Fig. 3B). These results confirm that targeting of VP1 antigen by Co1-conjugation into M cells skewed immunity toward Th2 response with increased IgA secretion because IL-6 is a major immune stimulator in tolerogenic mucosal environment and because IL-4 in LP is closely related with plasma cell differentiation (5,12).
Oral priming with Co1 ligand-conjugated VP1 of FMDV evoked the VP1-specific systemic immune response but not systemic tolerance induction
It is generally believed that tolerogenic immune response dominantly occurs against luminal antigens in the mucosal compartment and is one of the major obstacles in developing effective oral mucosal vaccines (14). In order to confirm whether oral administration of Co1-conjugated VP1 induces efficient systemic immunity against VP1 without tolerance induction, we measured the IgG SCs level in SPLs. The number of VP1-specific IgG SCs in SPLs prepared from the mice orally primed with VP1-Co1 and then challenged with VP1 alone was about more than 8 times higher than that of the mice fed VP1 alone (Fig. 4A). The increased level of VP1-specific IgG SCs was turn out to be due to Th2-skewed immune response against VP1 in that oral administration of VP1-Co1 significantly (p<0.05) increased the number of Th2-type cytokine (IL-4 and IL-5) SCs to a level about 2-fold higher than that with VP1 alone, although the level of Th1 type cytokine (IFN-γ) SCs was not changed (data not shown, Fig. 4B). In addition, corresponding enhancement of VP1-specific lymphocyte priming in the systemic compartment without tolerance induction by oral administration of Co1A-conjugated VP1 was verified in lymphocyte proliferation assay after in vitro stimulation with the VP1 antigen (Fig. 4C). Collectively, these results suggest that oral priming with Co1A-conjugated antigen may enhance systemic immune response without tolerance induction through T cell-mediated antigen-specific IgG production.
DISCUSSION
Current FMDV vaccine is an injection-based chemically inactivated vaccine and many disadvantages have been exposed such as the need for cold-chain storage, the risk of virus release during vaccine production, and the difficulty in serological distinction between infected and vaccinated animals (15). On the contrary, oral mucosal vaccine against FMDV infection using recombinant antigen may avoid the disadvantages of injection-based vaccine. Moreover, the suggestion by recent studies that induction of mucosal IgA is correlated with protection in FMDV-infected pigs indicates the need for developing oral mucosal vaccine against FMDV infection (4). SIgA is a unique effector element in the mucosal immune compartment and regulation for SIgA generation is critical in the success and quality of antigen-specific mucosal immune response induction (16). SIgA plays a role in the first line of protection through immune exclusion, intracellular neutralization, and antigen excretion. Role of SIgA in immune exclusion is mediated through the entrapment of antigen or bacteria and the inhibition of their interaction with epithelial cells (17). The presence of SIgA in endosome vesicles can also neutralize the replication of viruses (e.g., Sendai virus and rotavirus) as well as the activity of toxins. For example, in rotavirus, non-neutralizing IgA to VP6 can inhibit chronic murine rotavirus infection through its interactions with the virus in the luminal side of the intestinal tract (16). SIgA can also excrete luminal antigens by endocytosis of the antigen-dimeric IgA-J chain-pIgR complex. A recent study showed HIV virions excreted from the basolateral side to the apical area of the epithelial cells after interacting with IgA monoclonal antibodies and pIgR (18).
Introduction of vaccine materials into mucosal lymphoid tissue is essentially required to generate antigen-specific SIgA response (19). In mucosal epithelium, influx of luminal antigens was limited by pathways involving M cells, goblet cells, and specific dendritic cells, although transcytosis of macromolecules mainly occurs in M cells through the non-specific or specific receptor-mediated process (20). Therefore, research interest has been concentrated on elaboration of strategies to target antigens to M cells for developing effective oral mucosal vaccines. For example, M cell-specific antibody effectively delivers the conjugated antigen into the M cells, and UEA-1 interacting with α-L-fucose on M cells is used as a delivery mediator (13). Our previous studies also suggested that M cell-targeting ligand Co1 interacts with C5aR expressed on M cells and that this interaction is related with antigen delivery and induction of antigen-specific immune response (9,10). Our study also showed that Co1-conjugated viral antigen VP1 of FMDV more effectively interacted with M cells in ileal PPs compared to VP1 alone without Co1 ligand conjugation (Fig. 1). In addition, oral priming with VP1-Co1 enhanced antigen-specific IgA and IgG responses (Fig. 2 and 4A). In fact, generation of antigen-specific Ig is closely related with T cell-dependent reaction mediated by specific cytokines. For example, IL-4 helps isotype switching to IgA, and IL-5 which is predominantly expressed by T cells allows for proliferation and differentiation of pre-activated B cells (19). In addition, IL-6 plays multiple regulator roles in enhancing immune stimulation and IgA secretion (5). As shown in previous studies, this study also showed that Co1-mediated immune induction was due to Th2-skewed immune stimulation in systemic and mucosal compartments, and this pattern of immune stimulation supported the observed enhanced Ig response without tolerance induction (Fig. 3 and 4B). Although we did not monitor the neutralizing and/or protective ability of VP1-specific immunity induced by oral administration of Co1-conjugated VP1 in this study due to restriction in the experiments using FMDV, we believe the platform for the development of effective oral mucosal vaccine against FMDV infection in the future has been provided by this study.
In summary, we conjugated the M cell-targeting ligand, Co1, to a practical pathogenic antigen, VP1 of FMDV, and monitored the effects of ligand conjugation on the induction of immune response after oral mucosal priming and systemic challenge with the antigen. Effective delivery of VPI into the M cells of PP was confirmed through in vitro binding experiments and efficient VP1-specific immune response induction both in mucosal and systemic compartments was confirmed after oral administration of the antigen to mice. Collectively, we suggest that the M cell-targeting ligand Co1 may be useful as a mucosal vaccine adjuvant in FMDV oral mucosal vaccine development through effective delivery of the ligand-conjugated antigen into the mucosal immune compartment and efficient induction of mucosal and systemic immune responses.


 XML Download
XML Download