

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The adjuvant activity of alum in diphtheria toxoid immunization was first reported in 1926 (1) and it is the most widely used adjuvant in human vaccines. Nonetheless, it is surprising that the mechanism of its adjuvanticity still remains elusive. Alum was originally proposed to ensure a long-lasting immune response through formation of a depot. However, it has been shown that the depot of alum is not necessary for adjuvanticity (2,3). It is relatively well accepted that alum predominantly induces humoral immunity, in particular IgG1 response. It has been shown that alum adjuvant boosts IgG1 response by inducing uric acid and activating inflammatory DCs (4). Kool et al. have also demonstrated that alum stimulates inflammatory DCs through activation of NOD-like receptor family, pyrin domain containing 3 (NLRP3) inflammasome (5). Further, NLRP3 mediates Ag-specific Ab response and Th2-dependent inflammation induced by alum (6,7). However, the involvement of NLRP3 in the Th2 immune response to alum adjuvants is controversial. Thus, NLRP3 inflammasome is dispensable for aluminum adjuvanticity (8,9) and not to play a role in Th2-mediated immune responses in the airway (10). The discrepancy in results obtained by different group can not be resolved at the moment. Other immune cells are also known to be involved in alum-stimulated Ab enhancement: eosinophils (11) and NKT cells (12). However, there were no reports dealing the direct effect of alum on B cells. We assumed that alum may have effect on B cells since all kinds of TLRs and NLRs are expressed in B cells (13). The present study explored whether alum can regulate murine B cells-antibody synthesis. We found that alum can directly modulate mouse B cells to produce IgG1 Ab.
Go to : 
MATERIALS AND METHODS
Mice
BALB/c mice were purchased from Daehan Biolink Co. (Seoul, Korea). They were maintained on an 8:16-h light: dark cycle in an animal environmental control chamber (Daehan Biolink. Co., Korea). Animal care was in accordance with the institutional guidelines of Kangwon National University. Eight- to twelve-week-old female mice were used in this study.
Reagents
Imject Alum (hereafter simply called alum) was from Pierce Biotechnology (Rockford, IL, USA). IL-4 and TGF-β1 were purchased from R&D Systems (Minneapolis, MN, USA). LPS (Escherichia coli O111:B4) was from Sigma Chemical Co. (St. Louis, MO, USA). TRIZOL reagent was purchased from Invitrogen Life Technologies (Carlsbad, CA, USA). The antibodies used in ELISA were purchased from Southern Biotechnology (Birmingham, AL, USA). Anti-mouse CD3ε Ab and anti-mouse CD28 Ab were purchased from BD Biosciences (San Jose, CA, USA).
Cell culture
The murine B cell lymphoma line, CH12F3-2A (surface µ+) (14) was provided by Dr. T. Honjo (Kyoto University, Japan). The cells were cultured in RPMI-1640 (100 U/ml penicillin; 100µg/ml streptomycin) plus 10% fetal bovine serum (HyClone Labs, Logan, UT, USA) in a humidified CO2 incubator. Mouse spleen B cell population was prepared as described before (15). A total of 2×106 cells/well were cultured in flat-bottomed, 24-well tissue culture plates (SPL, Korea) in a volume of 2 ml complete medium or a total of 2×105 cells/well were cultured in flat-bottomed, 96-well tissue culture plates in a volume of 200µl complete medium with added stimulants.
Plasmid and transfection
AID promoter reporter was given by Dr. M. Sugai (Center for Molecular Biology and Genetics, Kyoto University) (16). Transfection was performed by electroporation with a Gene Pulser II (Bio-Rad, USA) as described (15). Reporter plasmids were cotransfected with expression plasmids and pCMV-βgal (Stratagene), and luciferase and β-gal assays were performed as described (15).
RNA preparation and RT-PCR
RNA preparation, reverse transcription, and PCR were performed as described before (15). PCR primers were synthesized by Bioneer Corp. (Seoul, Korea): GLTγ1 sense, 5'-CAG CCT GGT GTC AAC TAG-3' and antisense, 5'-CTG TAC ATA TGC AAG GCT-3' (product size : 532 bp); GLTγ2a sense, 5'-GCT GAT GTA CCT ACC TGA GAG A-3' and antisense, 5'-GCT GGG CCA GGT GCT CGA GGT T-3' (product size : 394 bp); GLTγ2b sense, 5'-GGG AGA GCA CTG GGC CTT-3' and antisense, 5'-AGT CAC TGA CTC AGG GAA-3' (product size : 318 bp); GLTγ3 sense, 5'- CAA GTG GAT CTG AAC ACA-3' and antisense, 5'-GGC TCC ATA GTT CCA TT-3' (product size : 349 bp); GLTα sense, 5'-CAA GAA GGA GAA GGT GAT TCA G-3' and antisense, 5'-GAG CTG GTG GGA GTG TCA GTG-3' (product size : 206 bp); CTγ1 sense, 5'-CCA AAA CAG GAA CAG AGA CG-3' and antisense, 5'-GTT CCA GGT CAC TGT CA-3' (product size : 434 bp); AID sense, 5'-TGC TAC GTG GTG AAG AGG AG-3' and antisense, 5'-TCC CAG TCT GAG ATG TAG CG-3' (product size : 119 bp); IFN-γ sense, 5'-ACA CTG CAT CTT GGC TTT GC-3' and antisense, 5'-TGG ACC TGT GGG TTG TTG AC-3' (product size : 369 bp); T-bet sense, 5'-ACC AAC AAC AAG GGG GCT TC-3' and antisense, 5'-CTC TGG CTC TCC ATC ATT CAC C-3' (product size : 111 bp); IL-4 sense, 5'-ATA TCC ACG GAT GCG ACA AA-3' and antisense, 5'-AAG CCC GAA AGA GTC TCT GC-3' (product size : 252 bp); GATA-3 sense, 5'-ACA GAA GGC AGG GAG TGT GTG AAC-3' and antisense, 5'-TTT TAT GGT AGA GTC CGC AGG C-3' (product size : 118 bp); CXCR5 sense, 5'-GAC CTT CAA CCG TGC CTT TCT C-3' and antisense, 5'-GAA CTT GCC CTC AGT CTG TAA TCC-3' (product size : 139 bp); Foxp3 sense, 5'-CTT CAT GCA TCA GCT CTC CA-3' and antisense, 5'-AGA CTC CAT TTG CCA GCA GT-3' (product size : 293 bp); β-actin sense, 5'-CAT GTT TGA GAC CTT CAA CAC CCC-3' and antisense, 5'-GCC ATC TCC TGC TCG AAG TCT AG-3' (product size : 320 bp). PCR reactions for β-actin were performed in parallel in order to normalize cDNA concentrations within each set of samples. PCR products were separated on a 2% agarose gel and photographed.
Statistical analysis
Statistical differences between experimental groups were determined by analyses of variance, and values of p<0.05 by unpaired two-tailed Student's t-test were considered significant.
Go to :
RESULTS AND DISCUSSION
Effect of alum on the expression of Ig isotypes in mouse B cells
We first explore whether alum affects Ig production by LPS-stimulated mouse B cells. As shown in Fig. 1, alum consistently increased IgG1 isotype secretion in a dose-dependent manner. In this, though less potent, alum also increased secretion of IgG2b and IgG3 isotypes. Thus, potency of alum on IgG isotypes appeared to be IgG1>IgG3>IgG2b. In contrast, it little affected secretion of IgM, IgG2a and IgA isotypes. In the absence of LPS, alum by itself did not result in secretion of any isotypes (data not shown). These results clearly show that alum directly modulates B cells to produce IgG1 isotype under the influence of LPS, a mouse B cell polyclonal activator. This is a somewhat surprising finding since no any reports have demonstrated the direct effect of alum on Ig synthesis by B cells ever since its adjuvanticity was discovered (1).
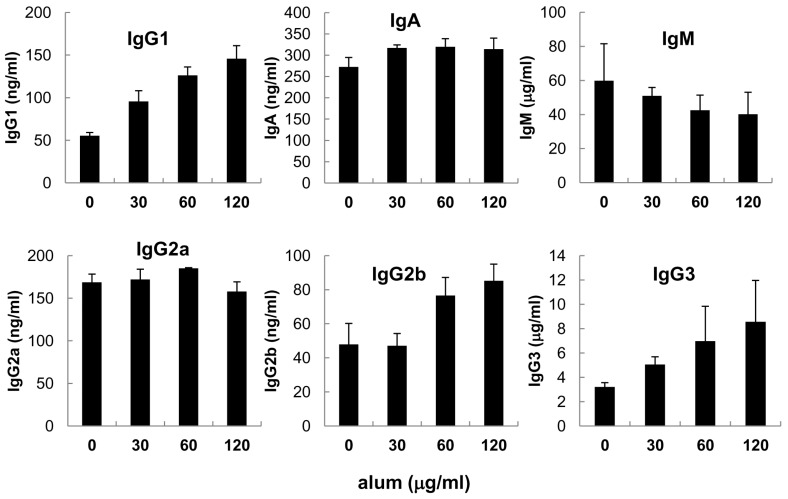 | Figure 1Effect of alum on Igs secretion in mouse spleen B cells. Mouse spleen B cells were cultured with LPS (12.5µg/ml) and alum (30µg/ml, 60µg/ml, 120µg/ml). After 7 days of culture, supernatants were collected and Igs productions were determined by isotype-specific ELISA. Data are means of triplicate samples± SEM.
|
Effect of alum on the viability of mouse spleen B cells
Since alum markedly influenced B cells in the synthesis of Ig isotypes, it was necessary to determine its effect on B cell growth. In the absence of LPS, alum did not increase cell viability but alum at the high dose (120µg/ml) delayed cell death (Fig. 2). In contrast, its effect was marginal in the presence of LPS. These results indicate that the modulatory effect of alum on Ig synthesis is not associated with its potential effect on B cell growth.

Effect of alum on IL-4-induced IgG1 and TGF-β1-induced IgA isotype expression
IL-4 and TGF-β1 are well established switching factors for IgG1 and IgA isotypes, respectively (17,18). It was curious how alum affects production of IL-4-induced IgG1 and TGF-β1-induced IgA isotypes. As shown in Fig. 3, IL-4 increased IgG1 production more strongly than alum. However, IL-4 and alum in combination did not result in any additive effect on IgG1 production. Similary, TGF-β1-induced IgA production was not further augmented by treatment of alum. These results suggest that signalling pathway of IL-4 and alum toward IgG1 gene expression is identical or at least largely overlapping, of which mechanisms remains to be determined.
 | Figure 3Effect of alum on Ig secretion in the presence of IL-4 and TGF-β1. Mouse spleen B cells were cultured with LPS (12.5µg/ml), alum (120µg/ml), IL-4 (10 ng/ml), and TGF-β1 (0.5 ng/ml) for 7 days. Culture supernatants were collected and Igs productions were determined by isotype-specific ELISA. Data are means of triplicate samples±SEM. *p<0.05.
|
Alum increases expression of IgG1 germ line transcripts, an indicative of IgG1 isotype switching
Because alum directly modulates B cells to differentiate mainly into IgG1 B cells, it was necessary to investigate if alum increases IgG1 production by increasing the total number of cells that secrete IgG1, or simply by increasing the amount of IgG1 secreted per cell. As shown in Fig. 4, there was an increase in the number of IgG1-secreting cells in alum-stimulated cultures. This was paralleled by an increase in the total amount of IgG1 secreted (Fig. 1). Here, IgG1-spot sizes between cultures of LPS alone and LPS plus alum were not different, indicating that alum does not increase the amount of IgG1 secreted per cell.
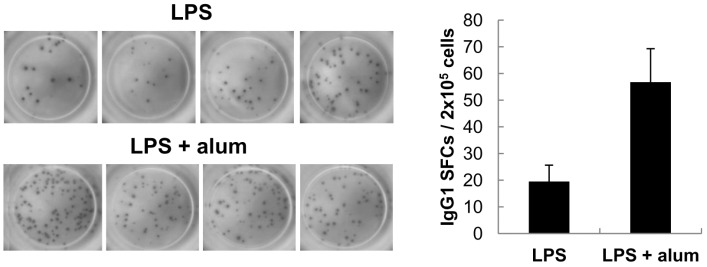
Increases in the number of IgG1-secreting cells in the above experiments is: 1) due to an increase in the frequency of B cells that switch to express the IgG1 isotype after stimulation with alum, or 2) due to increased proliferation of B cells that are already committed to express the IgG1 isotype. To distinguish between these two possibilities, we explored whether alum affects IgG1 class switch recombination (CSR). Transcription of unrearranged Cγ1 gene to produce germ-line transcript γ1 (GLTγ1) precedes CSR to IgG1 (Fig. 5A). Once IgG1 CSR occurs, post-switch γ1 transcripts (PSTγ1) and γ1 circle transcript (CTγ1) are also expressed. Therefore, expression of GLTγ1, CTγ1 and PSTγ1, can be used to indicate active IgG1 CSR (19-21). As shown in Fig. 5B, alum increased the expression of GLTγ1 and CTγ1 but not other GLTs. Since alum did not stimulate cell proliferation (Fig. 2), it is regarded that alum induces IgG1 production mainly through IgG1 CSR.
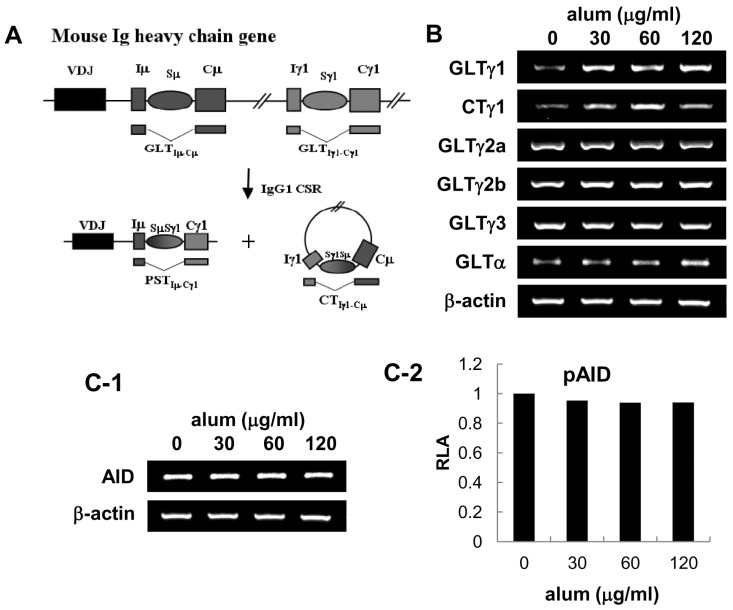 | Figure 5Effect of alum on the expression of Ig GLTs and AID. Mouse spleen B cells were cultured with LPS (12.5µg/ml) and alum as indicated for 2 days. (A) Diagram of DNA recombination occurring during switching to IgG1. Rectangles and ovals represent exons and S regions, respectively. RNA transcripts are indicated beneath the DNA diagrams. (B) Levels of GLTs, CTγ1, and β-actin were measured by RT-PCR. (C-1) Effect of alum on endogenous AID transcriptional level was measured after 2 days of culture by RT-PCR. (C-2) CH12F3-2A cells were transfected with 5µg of pAID1. Luciferase activity was determined following alum treatment as indicated for 48 h. Transfection efficiency was normalized to β-gal activities. Data represent average of two independent samples.
|
Alum neither increases activation-induced cytidine deaminase (AID) nor activates Th2 cell differentiation
It is well established that AID is prerequisite for Ig CSR (19,22), it was important to ask if alum has effect on AID expression. As shown in Fig. 5C, alum did not increase both the expression of endogenous AID transcripts and the exogenous AID promoter activity, suggesting that alum does not cause general Ig CSR. Finally, we investigated whether alum directly regulate CD4 T cell differentiation. To assess this issue, we measured the transcriptional levels of canonical markers for each T cell subsets: IFN-γ and T-bet for Th1, IL-4 and GATA-3 for Th2, CXCR5 for follicular helper T cell, Foxp3 for regulatory T cell. We found no significant change in CD4 T cell subset population under the influence of alum (Fig. 6). Again, these results support our finding that alum by itself can act on B cells resulting in IgG1 CSR without any helper T cell subset including Th2. Then, what would be the mechanisms by which alum directs B cells for Ig regulation? In this regard, one needs to consider at least three possible ways which were the case in DCs. That is, alum may direct B cells 1) through release of DNA (23), 2) through forming of inflammasome (6,7), 3) through lipid rafts (24).
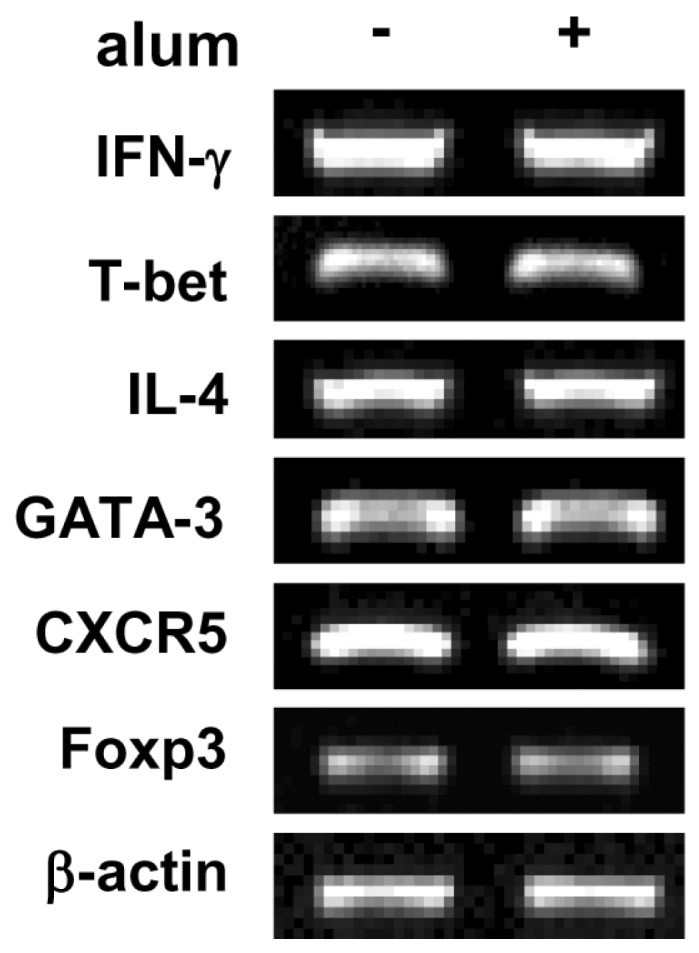 | Figure 6Effect of alum on T cell differentiation. Mouse spleen whole cells were stimulated with plate bound anti-CD3 mAb, soluble anti-CD28 mAb and IL-2. Spleen whole cells were incubated with alum (120µg/ml) for 2 days. Levels of IFN-γ, T-bet, IL-4, GATA-3, CXCR5, and Foxp3 transcripts were determined by RT-PCR. Fold increases represent relative DNA level normalized with the expression of β-actin cDNA.
|
Conclusion remarks
The present study demonstrates that alum can directly modulate B cells to commit IgG1 isotype switching. This implies that other immune cells such as dendritic cells, macrophages, and T helper cells are dispensable for the enhancement of IgG1 production by alum, though their involvement can not be formally excluded and to be elucidated in the future. Nevertheless, our findings of alum's direct effect on B cells would be useful to improve its vaccine adjuvanticity.
Go to :
 XML Download
XML Download