

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Vitamin C is regarded as an essential nutrient for the host defense mechanisms through the maintenance of immune homeostasis (1,2). High concentration of vitamin C is accumulated in most immune cells including, neutrophils, macrophages, B cells and T cells, and it is rapidly decreased by infection or stress (3-6). This suggests that vitamin C acts diversely on innate immunity as well as adoptive immunity. In addition, vitamin C enhances the proliferation and migration of lymphocytes as well as the killing activity of natural killer cells (7-10). The phagocytic activity of neutrophils and macrophages is also increased by vitamin C, but the number of T cells is not altered by vitamin C (5,11). Interestingly, the decrease of delayed type hypersensitivity (DTH) to several antigens in human is caused by vitamin C insufficiency, but it is normalized by the supplementation of the sufficient amount of vitamin C (9).
It is known that vitamin C enhances Th1 immune response via the stimulation of the production of IFN-γ and TNF-α. In fact, the decreased Th1 immune response against Helicobacter pylori infection was found in vitamin C insufficient Gulo (-/-) mice, which cannot synthesize vitamin C like a human due to the lack of L-gulono-γ-lactone oxidase (12). In accordance with the up-regulation of Th1 immune responses by vitamin C, the levels of IgG1 and IgE, Th2-related isotypes of antibody, were decreased, but IgG2C, Th1-related isotype of antibody, was increased in DTH animal models (13). However, its related specific mechanism of vitamin C is still largely unknown.
Dendritic cell (DC) is known as the most potent antigen presenting cell (APC). It can readily stimulate naïve T cells, since the essential co-stimulatory molecules, such as CD80 and CD86, are highly and constitutively expressed on its surface. Immature DCs can uptake antigens and migrate to lymph nodes (13). Then, in the presence of endogenous or exogenous inflammatory signals, DCs undergo maturation (14). Maturation of DCs is related with enhanced expression of co-stimulatory molecules such as CD80 and CD86, and MHC molecules. Several kinds of pattern recognition receptor (PRR) including toll like receptor (TLR) are closely involved in the maturation and activation of dendritic cells by the stimulation of danger associated molecular pattern (DAMP) (15,16). In addition, impaired production of reactive oxygen species (ROS) during Mycobacterium tuberculosis infection affects the differentiation of DCs (17). This suggests that anti-oxidant molecules, such as vitamin C and vitamin E, could modulate the activation and differentiation of DCs.
In the present study, we investigated whether vitamin C could modulate the activation of DCs through the regulation of CD80, CD86 and MHC II expression and its related mechanisms.
MATERIALS AND METHODS
Cells
Dendritic cell line, DC-1 cells were cultured in RPMI 1640 media containing 2 mM of L-glutamine, 100 units/ml of penicillin, 100 mg/ml of streptomycin, and 10% fetal bovine serum (Gibco, USA). Cells were cultured at 37℃ in a humidified atmosphere containing 5% CO2.
Detection of apoptosis
After cells (2×106) were exposed to various concentration of vitamin C (0.5, 1, 2 and 5 mM) for 24 hrs, they were collected and washed twice with cold PBS. Then they were resuspended in 1 × binding buffer at a concentration of 1×106 cells/ml. Cells were then incubated with 5µl of FITC conjugated Annexin V (BD Pharmingen, San Diego, CA, USA) at room temperature for 15 min in the dark. One microliter of 7-AAD (BD Pharmingen, San Diego, CA, USA) was added prior to flow cytometric analysis by FACSCaliber (BD Pharmingen, San Diego, CA, USA).
Measurement of reactive oxygen species (ROS)
Cells (5×104) were incubated in a 96-well plate. Cells were then further incubated with 50µM of 2',7'-dichlorofluorescein diacetate (DCFH-DA; Eastman Kodak, Rochester, NY, USA) at 37℃. After incubation, cells were analyzed with a Cytofluor 2350 plate reader (Millipore, Bedford, MA, USA) with excitation at 485 nm and emission at 525 nm.
Detection of CD80, CD86 and MHC II expression
Cells (4×105) were incubated for 24 hrs in the presence of 2.5 mM of vitamin C. And then, cells were stained with FITC-conjugated antibodies against CD80, CD86, and MHC class II (BD Pharmingen, USA) on ice for 30 min. The expression levels were analyzed using flow cytometric analysis by FACSCaliber (BD Pharmingen, San Diego, CA, USA). To determine the signaling pathway by vitamin C, cells were pre-treated with specific inhibitors, 10µM of SB203580, 20µM of LY294002, and 20µ M of SP600125 for 30 min, prior to exposure to 2.5 mM of vitamin C. Then the expression of CD80, CD86 and MHC class II were analyzed using flow cytometric analysis.
Western blot analysis
The phosphorylation of p38MAPK in DC1 upon the treatment of vitamin C was examined by western blot analysis. Cells were lysed in cell lysis buffer containing 1% Triton X-100 (v/v) in 20mM Tris-HCl, pH 8.3, 150 mM NaCl and protease and phosphatase inhibitor cocktail. Cytosolic proteins were quantified using bicinchoninic (BCA) protein assay, then 30µg of samples were loaded to 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) which were then transferred onto nitrocellulose membranes. The membranes were blocked in a solution of 5% skim milk in TBS-T buffer (20 mM Tris-HCl, pH 7.4, 150 mM NaCl and 0.1% Tween-20). Blocked membranes were incubated with rabbit anti-mouse p38MAPK (1:1,000) or rabbit anti-mouse phosphor-p38MAPK (1:1,000) (Cell Signaling Technology, USA). Horseradish peroxidase-conjugated donkey anti-rabbit IgG (1:2,000) (Santa Cruz Technology, USA) was used secondary anti-body. Immunodetected bands were visualized with the ECL system (Amersham Pharmacia Biotech, UK), and results were analyzed using densitometer program (Scion image).
RESULTS
Vitamin C acts as an effective anti-oxidant on DC1
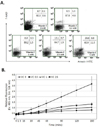
To determine the optimal concentration of vitamin C without cytotoxicity, we examine the induction of apoptosis on DC-1 after treatment of 0, 0.5, 1, 2.5 and 5 mM of vitamin C. As shown in Fig. 1A, more than 85% of cells were viable, even with the treatment of 2.5 mM of vitamin C. However, 60.9% of cells were viable by the treatment of 5 mM of vitamin C (Fig. 1A). Next, we examined intracellular ROS levels in DC-1 after treatment of vitamin C. Without treatment of vitamin C, intracellular ROS levels were spontaneously increased in DC-1, but it was suppressed by the treatment of vitamin C. However, there were no differences on intracellular ROS levels, among the groups that exposed vitamin C (Fig. 1B). Therefore, the relatively high concentration of vitamin C did not induce apoptosis on DC-1. And it effectively acts as an anti-oxidant molecule.
Vitamin C up-regulates the expression of CD80, CD86 and MHC class II on DC-1
It is generally known that oxidative stress was suppresses the immune responses (18). However, we have already shown that vitamin C effectively suppresses the increase of ROS levels in DC-1. Therefore, we next compared the expression levels of CD80, CD86m and MHC class II molecules on DC-1 at 24 hrs after treatment of 1 and 2.5 mM of vitamin C. Even though we could not induce those three of molecules on DC-1 cells by the treatment of 1 mM of vitamin C, we found the extensive induction of CD80, CD86 and MHC class II on DC-1 cells by the treatment of 2.5 mM of vitamin C (Fig. 2). When we considered that DC-1 shows characteristics of immature DCs, it implies that vitamin C plays an important role in the maturation of immature dendritic cells through the up-regulation of CD80, CD86 and MHC class II molecules.
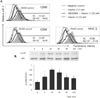
The up-regulation of CD80, CD86 and MHC class II on DC-1 by the treatment of vitamin C is mediated by the activation of p38 MAPK
Finally, we investigated which kind of signal transducing molecule is involved in the expression of CD80, CD86 and MHC class II on DC-1 by the treatment of vitamin C. Even though it is in cancer cells, we found that vitamin C regulates the activation of p38MAPK in our previous study (19). Therefore, the changing on the expression of those three molecules on DC-1 by the treatment of inhibitor for p38MAPK, SB203580, prior to the treatment of vitamin C was examined. As shown in Fig. 3A, we found that the expressions of CD80, CD86 and MHC class II on DC-1 were definitely decreased by the pre-treatment of SB203580 (Fig. 3A). We confirmed whether vitamin C could induce the phosphorylation of p38MAPK in DC-1 by immunoblotting. As shown in Fig. 3B, the phosphorylation of p38MAPK was increased by the treatment of 2.5 mM of vitamin C. Taken together, vitamin C up-regulates CD80, CD86 and MHC class II on DC-1 via the activation of p38MAPK.
DISCUSSION
Vitamin C effectively prevents oxidative damages through the scavenging of oxygen radicals. The physiological vitamin C concentration in human serum is known as 70~85µM (20,21). Serum concentration of vitamin C is determined by the route of administration. According to the report by Sebastian J et al, when 3 grams of vitamin C is administered via intravenous injection, concentration of vitamin C in serum is reached at 1,700µM, but administered via oral route; it is reached just 220µM (22). Even though vitamin C used in our experiment is 1 and 2.5 mM and it is much higher than its physiological concentration, serum concentration could be increased by the administration via intravenous injection of vitamin C. When we consider that most of recent trials of anti-cancer therapy by vitamin C is done by intravenous injection, it seems that the combination approach of vitamin C and chemotherapy or radiotherapy will give us the new insight of cancer therapy.
It is reported that the relatively high doses of vitamin C (more than 5 mM) induce the apoptosis on tumor cells. When B16F10 melanoma cells were exposed to vitamin C, vitamin C induces the decrease of mitochondrial membrane potential followed by the release of cytochrome C (23). In addition, it inhibited the uptake of iron, an essential factor for the survival of tumor cells, through the down-regulation of CD71 (transferrin receptor) expression (24). The relatively low doses of vitamin C (less than 1 mM) inhibit proliferation of tumor cells via the regulation of Chk2-p53-p21Waf1/Cip1 pathway and the expression of receptor for growth factor, such as insulin-like growth factor (IGF)-1 (25,26). Moreover, vitamin C could regulate angiogenic process of tumor through the down-regulation of vascular endothelial growth factor (VEGF) production. In this process, the expression of cyclooxygenase-2 (COX-2), a well-known pro-inflammatory mediator, was also down-regulated by vitamin C (27). Even though there are many reports regarding the direct role of vitamin C as an anti-tumor agent, the specific action of vitamin C as an immune regulator is largely unknown. We have previously reported that the surface expression of CD95 (Fas) on human stomach cancer cell line, SNU-1 was induced by the treatment of vitamin C (28). It suggests that the immune susceptibility of tumor could be increased by vitamin C. Since dendritic cell is the most potent APC to generate tumor specific T cells, we investigated the role of vitamin C on the maturation of dendritic cell in the present study.
In general, the expression of co-stimulatory molecules, CD80 and CD86, on DCs are known as a crucial secondary signal for the generation of effector T cells. Therefore, the up-regulation of CD80 and CD86 implies the enhancement of activity of DCs. According to the report by Iijima N et al., CD80 and CD86 expression of DCs was up-regulated by the treatment of tumor necrosis factor (TNF)-α (29). It is also reported that concanavalin A (Con A) and lipopolysaccharide (LPS) increases CD80 and CD86 expression on DCs at inflamed sites (30,31). Moreover, it is increased on monocytes in the patients with toxoplasma gondii infections to enhance proliferation of resting T cells in response to parasite-infected cells (32). It suggests that CD80 and CD86 play a crucial role in the initiation and maintenance of an immune response. To the point of this view, constitutively enhanced expressions of CD80 and CD86 by the treatment of vitamin C shown in the present study have the beneficial effect of the maintenance of immune responses. However, the expression of CD80 and CD86 was not up-regulated by the treatment of two of the most potent anti-oxidant, N-acetyl-l-cysteine (NAC) and reduced glutathione (GSH) (29). It seems that it is because of the different properties as an anti-oxidant between vitamin C and NAC and GSH. Vitamin C could be rapidly converted to its oxidized form, dehydroascorbate (DHA). In turn, DHA could be also rapidly converted to its reduced form, ascrobate (vitamin C). In contrast, NAC and GSH did not have redox cycle like vitamin C, even though they are more potent than vitamin C. Moreover, the reduction of glutathione disulfide (GSSG) to GSH is dependent on the action of vitamin C as an electron donor (33). Therefore, the different roles of anti-oxidants on the induction of CD80 and CD86 on DCs should be further investigated.
We have previously reported that vitamin C shows its activity through the regulation of p38MAPK and P42/44MAPK (19,25,27). In the present study, vitamin C regulates the activation of p38MAPK on DC-1. In addition, we found that vitamin C also modulates the activation of phsphoinositide-3 kinase (PI3K)/Akt in hepatocyte (our unpublished data). It might be that vitamin C shows its action after being transported into the cells through specific channel proteins. The reduced form of vitamin C, ascorbate is transported by sodium-dependent vitamin C transporter (SVCT)-1/2, and the oxidized form of vitamin C, dehydroascorbate (DHA) is transported by glucose transporter (GLUT) (34-36). That is to say, the phosphorylation of kinases by vitamin C is not mediated by receptor associated adaptor proteins, since SVCT-1/2 and GLUT are not receptor proteins, but channel proteins. Since we have shown that the intracellular ROS level was not decreased, when DC-1 cells were cultured medium without sodium, it seems that the activation of kinases is dependent on the activity of SVCT-1/2. To examine the mechanisms and its related molecules on the regulation of SVCT-1/2 activity on immune cells, including dendritic cells, we are now investigating with L-gulono-γ-lactone oxidase (Gulo) knock-out mice, which cannot synthesize vitamin C like a human (37).

 XML Download
XML Download