

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The development of anti-tumor immune therapies through the activation of antigen-specific immune function has the advantage that the side effects can be significantly reduced and the treatment effect of personalized therapy is excellent. Among these, dendritic cells (DCs) have been studied as anti-tumor vaccine since the late 1990's (1). DCs are the most potent antigen-presenting cells (APCs) and are equipped with the necessary co-stimulatory, adhesion and histocompatibility molecules for initiation of a primary immune response (2-4). DC-based vaccination trials in human solid cancers have aimed to activate the immune system to recognize and destroy tumor cells. The safety and the feasibility of anti-cancer DC vaccines have been confirmed in several clinical trials (5-9). Nevertheless, reports on DC therapy for lung cancer patients were limited (10-12).
Lung cancer is a leading cause of cancer-related death in both men and women worldwide (13). Although various therapeutic modalities, including surgical resection, radiation therapy, and chemotherapy, have been developed, the disease is rarely curable and the prognosis is dismal, with an overall 5-year survival rate of only 15% (14). Therefore the development of effective therapies is clearly needed for treatment of lung cancer. In our previous study (15), we verified the possibility of anti-tumor effect of cultured DCs as an adjuvant therapy for minimal residual disease state of lung cancer.
Although feasible anti-tumor effect of monocyte-derived DC (MoDC) vaccine was resulted in our previous study (15), antigen specific immune responses were weak with partial blocking of tumor growth. To improve the efficiency of anti-tumor effect using DC vaccine for lung cancer, we tested CD34+ hematopoietic stem cell derived DC (SDC) as an alternative in mouse model in this study. SDC can be applied in many clinical researches because hematopoietic stem cells generate a large number of mature DCs in vitro and able to differentiate into various stages. Ward et al. (16) reported that CD34+ DC generated with GM-CSF, IL-4, Flt-3L and SCF, capable of inducing naive T-cell responses and can be expanded to clinically useful numbers. There are reports that CD34+ DCs from core blood have anti-tumor activity (17-19). Our SDC were generated from CD34+-mouse bone marrow cell and compared to MoDC for their DC characters and in vivo anti-tumor effect.
MATERIALS AND METHODS
Animal
Specific pathogen-free female C57BL/6 mice, 5~6 weeks old, were purchased from the Dae-Han BioLink (EumSung, Korea). The mice were provided with water and food, adlibitum and quarantined under 12 h light: 12 h dark photoperiod in the animal care facility of the Samsung Biomedical Research Institute, Samsung Medical Center, Seoul, Korea. Animal care was performed following the ILAR guideline. The mice were acclimated for at least one week before any experiments were conducted.
Reagents & cell lines
Complete medium (CM) consist of RPMI 1640 with 10% Fetal Bovine Serum, 2 mM glutamine and 100 U/ml penicillin plus 100µg/ml streptomycin (all from GIBCO laboratories, Grand Island, NY, USA). Following antibodies for flow cytometric phenotyping were purchased from BD (SanDiego, CA); fluorescein isothiocyanate (FITC)-or phycoerythrin (PE)-labeled monoclonal Abs for MHC I/II, CD11c, CD8a, CD54 and CCR7. ELISA sets for cytokines including IL-10, IL-12, TGF-β and IFN-γ was purchased from BD (Sunnyvale, CA). For isolation of hematopoietic stem cells, a lineage cell depletion kit was obtained from Miltenyi Biotech Inc (Bergisch Gladbach, Germany). C57BL/6 syngeneic Lewis Lung Carcinoma (LLC) cell line was purchased from American type culture collection (ATCC) (Rockville, MD, USA). The following hybridoma cell lines were also purchased from ATCC; GK1.5 for anti-L3T4, 53.672 for anti-Lyt-2, RA3 for anti-B220, and J11d for anti-B cells/neutrophils.
Ex vivo generation of DCs
Mononuclear cells (MNCs) from bone marrow were obtained from the tibia and femur of C57BL/6 mouse sacrificed by cervical dislocation. The viability of red blood cell (RBC) free-MNCs was routinely demonstrated to be over 95% by trypan blue exclusion. To separate the myeloid lineage cells, RBC free-MNCs were treated with antibodies against L3T4, Lyt-2, B220, and B cell/neutrophil with complement. Purified cells (1×106/ml) were incubated with GM-CSF and IL-4 (1×103 units/ml each) at 37℃ for 7 d in a humidified CO2 incubator for MoDC differentiation. To generate SDCs, CD34+ cells were isolated from the bone marrow cells by MACS lineage cell depletion kit and cultured with GM-CSF, SCF and IL-4 for 14 days. Eighteen hours before cell harvest, tumor cell lysate (50µg protein/ml) was added to culture media. Tumor cell lysate was prepared from cultured LLC cells by a freezing-thawing process that was repeated six times in liquid nitrogen (-180℃) and an incubator (37℃). Protein quantification of the tumor cell lysate was performed using centrifuged (1,500 rpm, 15 min) supernatant and the Bradford method (Bio-Rad Protein Assay; Bio-Rad Laboratories, Hercules, CA, USA). Cultured cells were characterized as therapeutic DCs by surface marker phenotyping and cytokine secretion into the culture supernatant.
Tumor inoculation and injection of therapeutic-DCs
Syngeneic LLC cells were inoculated intravenously into C57BL/6 mice on day 0. Starting from the day after the tumor cell injection, antigen pulsed therapeutic-DCs were injected twice by one week interval into the peritoneum of mice. This way the minimal residual disease status was simulated by having the tumor cells in the blood without established burdens. Each experimental group was composed of 5~7 mice and the control group was treated with saline. Animals were sacrificed at three weeks after the final DC injection to observe tumor growth in the lung and to analyze the systemic immune modulation. The treatment schedule is summarized in Fig. 1. Pulmonary tumor burden and IFN-γ secreting CD8+ T cell proportion (ELISPOT) were detected from the splenocytes of mice in each group.
Immunophenotypic analysis of cultured cells by flow cytometry
Phenotype of cultured cells was analyzed by direct immunefluorescence staining of cell surface antigens using FITC or PE conjugated antibodies against MHC I/II, CD11c, CD8a, CD54 and CCR7. Single cells of cultured DC were incubated with fluorescence-labeled surface antibodies in PBS with 0.1% sodium azide and 1% FBS (PBS-CS) for 40 min at 4℃. Within 1 h of the antibody labeling, cells in the 500µl PBS-CS were analyzed by flow cytometry (FACS Vantage; BD, CA, USA).
Measurement of cytokine secretion from DCs
The concentrations of IL-10, IL-12, TGF-β and IFN-γ in culture supernatant of DC were measured using commercial ELISA kits (BD, Sunnyvale, CA).
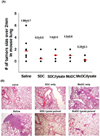
Mouse lung observation (In vivo anti-tumor effect of DC vaccine)
Three weeks after the last DC injection, tumor bearing mice were sacrificed. In order to investigate in vivo anti-tumor effect of DC vaccine, lung with heart of the mice were eliminated. To quantify the effect on the tumor establishment and growth, tumor lesions bigger than 2 mm were counted under a microscope (×100). Then these lungs were paraffin embedded and stained with H&E to observe the histo-pathological alterations. Histological observation was performed under the microscope (Leica DM 3000). The picture was presented with ×100 magnification.
Analysis of IFN-γ producing cells using enzyme-linked immunospot (ELISPOT) assay
The ELISPOT assay was adopted to detect and enumerate individual cells that secrete IFN-γ protein in vitro upon exposure to antigen. ELISPOT IFN-γ assay kit were purchased from AID (AID, Strassberg, Germany) and assays were performed according to the manufacturer's instruction. In brief, using magnetic bead cell separation system (MACS™ Miltenyi Biotec, Germany), CD3+ T cells were purified from the splenocytes (2×105 cells) and stimulated in vitro with tumor lysate on a 96-well pre-coated plate. The plate was incubated for 20 h at 37℃ with 5% CO2. After washing, each well was added with detection antibody and was incubated for 2 h at room temperature. The plate was incubated with alkaline phosphatase conjugate and developed with BCIP/NBT substrate solution. Visible spots were enumerated using an automated AID ELISPOT reader (AID, Strassberg, Germany) and the default program.
Statistical analysis
The analyzed values are reported as the mean value±standard error (SE). Each experimental group was composed of 5~7 mice, and the same experimental protocol was repeated at least two times with similar results. Statistical significance was determined by two-tailed Student's t-test. p-values<0.05 were interpreted to represent statistically significant differences.
RESULTS
Characterizations of generated DC
Generated therapeutic DCs were verified through cytokine secretion and phenotype expression. From the SDC supernatant, immune-stimulatory cytokine IL-12 was measured significantly higher level than MoDC (424.82±87.2 vs. 73.51±6.15 pg/ml for SDC vs. MoDC group, 574.38±191 vs. 66.06±2.38 pg/ml for SDC/lysate vs. MoDC/lysate group respectively) (Fig. 2). On the other hand, in the MoDC supernatant, IFN-γ as an immune-stimulatory cytokine were determined significantly higher level than SDC (399.79±105.59 vs. 2,601.31±680.13 pg/ml for SDC vs. MoDC group, 352.69±21.19 vs. 2,090.26±329.86 pg/ml for SDC/lysate vs. MoDC/lysate group respectively) (Fig. 2). Although secretion of immune-inhibitory cytokine IL-10 or TGF-β was observed from each DC group (IL-10 in MoDC group vs. TGF-β in SDC group), there were amounts of immune-stimulatory cytokines which are considered as counterbalance. Over 90% MoDC & SDC were expressed MHC I/II molecules (Fig. 3A) (The data of MHC I does not shown). However, a DC marker CD11c expression was prominent in SDC (Table I). The proportion of CD11c+CD8a+ cell, known as Th1-inducible DC subset was similar in two DC types (Table I) (Fig. 3B). Although the CD54 expression was similar in both DCs, CCR7 expression was prominent in MoDC (Table I) suggesting the better migratory function of MoDC (Fig. 3C). The characters of both SDC and MoDC revealed that both DCs could induce anti-tumor immunity with different mechanisms.
In vivo anti-tumor effect of DCs
To quantify the effect on the tumor establishment and growth, we eliminated lung with heart of the mice and observed tumor lesions under a microscope. After the DC treatment in LLC injected mice, countable tumor burdens (over than 2 mm under a ×100 microscope) were decreased compared to saline treated group (Fig. 4A). However, no statistical significance was found between the MoDC and SDC as well as with/without tumor lysate group. Despite the fact that statistically significant differences through eye observation were not found, histological alterations were observed from the H&E stained lung sections (Fig. 4B). In DC treated group, a significant amount of tumor was reduced. Moreover, SDC treated group pulsed with LLC lysate shows definite tumor regression. This H&E staining shows SDC/lysate induce more powerful anti-tumor immunity than MoDC/lysate in mouse lung cancer.
Ag-specific T cell immune response
To confirm SDC/lysate induce more systemic anti-tumor immunity than MoDC/lysate, we checked effector T cells in splenic lymphocytes from LLC bearing mice. To measure the anti-tumor effector T cells, ELISPOT assay counting IFN-γ secreting CD8+ T cells was operated (Fig. 5). The frequency of IFN-γ secreting CD8+ T cells in SDC treated group was significantly higher than that in MoDC treated group (58.7±8.3 vs. 11.7±1.8 spots for SDC/lysate vs. MoDC/lysate treated group, respectively). It shows SDC/lysate induce naïve T cells to effector T cells which lead to regress residual tumor in mouse lung. The therapeutic responses were associated with induction of IFN-γ secreting CD8+ T cells.
DISCUSSION
In spite of the aggressive treatment with surgery, radiation, and chemotherapy, the long-term survival for patients with lung cancer still remains low. Even patients with early stage disease often succumb to lung cancer due to the development of metastasis, indicating the need for effective approaches for the systemic therapy of this condition (20). Based on the antigen specificity of the immune system and the safety profile of cancer vaccines, the effective immunotherapy would be an ideal adjuvant, following initial clinical responses to definitive therapy (21). Even though there have been numerous clinical trials for various types of cancer, there are few DC vaccine trials in patients with lung cancer, and many aspects related to the immunotherapy are unknown. Our previous study suggests the possible anti-tumor effect of cultured DCs as an adjuvant therapy for minimal residual disease state of lung cancer (15). Nevertheless, MoDC vaccine in previous study shows weak antigen-specific immune response and incomplete tumor regression. In order to increase the efficiency of anti-tumor response using DC therapy, we introduced other culture methods as alternatives. Hematopoietic stem cell (HSC) as an origin of DC culture has advantage that the culture yield can be increased by more than 30 to 60 times depending on the culture protocol and/or incubation period. Also the HSC has the potential to differentiate into the various type of cell. Authors reported the HSC-derived DC vaccine effect on the renal cell carcinoma and breast cancer patients (7). No report for the effects of SDC on lung cancer treatment was found. The characters of SDC cultured in this protocol allow expectations that SDC may work to induce the anti-tumor immunity, since these cells produce significantly more IL-12 and lower IL-10 than MoDC (Fig. 2). Unlike SDC, cultured MoDC produced significantly higher IFN-γ and lower TGF-β than SDC (Fig. 2) which also may work as anti-tumor effector by different mechanism. Indeed, the proportion of cells expressing CD11c and CD8a, an immunogenic DC subset inducing Th1 response in mouse (22,23), was similar in both DC culture (Fig. 3) with similar response to inhibiting tumor growth (Fig. 4A). One day after inoculation of LLC cells, therapeutic-DCs were injected twice by one week interval into the peritoneum of tumor cell injected mice. Thus the effect of DC could be analyzed as an adjuvant treated after removing the tumor burden. Although DC vaccines inhibit the tumor growth in the lung, there were no statistical significances between the effect of MoDC and SDC when we observed tumor burden over 2 mm size under the microscope (Fig. 4A). This phenomenon was considered because we missed the presence of tumor under 2 mm size. For this reason, we observed lung tissue through H&E staining to determine histo-pathological changes (Fig. 4B). The entire DC treated group revealed the remarkable tumor regression. Affirmatively, no small tumorigenic changes other than countable tumors were observed in the tumor lysate- pulsed SDC treated lung. Also SDC significantly induced antigen-specific IFN-γ secreting CD8+ T cells than MoDC did (Fig. 5). Collectively, although both MoDC and SDC can induce the anti-tumor response, SDC considered as efficient inducer of anti-tumor immunity than MoDC in the mouse lung cancer.





 XML Download
XML Download