

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
α-Mangostin is a xanthone derivative included in the fruit hull of mangosteen (Garcinia mangostana L.), and recently it was reported that α-Mangostin induced apoptosis of several cancer cells via sarcoplasmic/endoplasmic Ca2+ ATPase (SAECA) inhibition and autophagy activation (1,2). Although α-Mangostin has an inhibitory effect on SAECA like thapsigargin, a sesquiterpene lactone, initially isolated from a plant, Thapsia garganica, it was different from thapsigargin in that α-Mangostin activated autophagy pathway, whereas thapsigargin induced autophagy arrest (3). Disruption of endoplasmic reticulum (ER) Ca2+ homeostasis which was mediated by the inhibition of SAECA, resulted in Ca2+ deprivation in ER, raised cytosolic Ca2+ concentration, and consequently apoptosis (4).
Autophagy, especially macroautophagy, is a lysosome-dependent degradation pathway for homeostatic regulation of cells which involved in degradation of cytosolic protein and organelles (5). Several studies have shown that ER stress induced autophagy flow via JNK activation or PERK/eIF2α phosphorylation (6,7). In the current study, the administration of α-Mangostin efficiently inhibited tumor growth in correlation with previous reports showing the anticancer activity of α-Mangostin. Although the role of ER stress in cancer has been studied in depth (8,9), the effect of ER stress-mediated autophagy under ER stress upon anticancer activity was not studied well. Thus we tried to find out the effect of ER stress with autophagy regulators, including α-Mangostin and thapsigargin, on antitumor activity. Interestingly, despite the similar inhibitory effect on SAECA, α-Mangostin did not increase ER stress in vitro, whereas α-Mangostin reduced ER stress induced by thapsigargin treatment. Collectively, these results suggested that autophagy activation by α-Mangostin could overcome ER stress mediated by Ca2+ ATPase blockade by itself. However, autophagy arrest by thapsigargin completely inhibited the antitumor effect by α-Mangostin, suggesting that autophagy activation in cancer might be an important strategy to achieve successful antitumor activity in the presence of ER stress.
MATERIALS AND METHODS
Mice
BALB/c mice and C57BL/6 mice (Orient-Bio Ltd, Charles River Laboratories, Seoul, Korea) were purchased at 6 weeks of age and used in all of the experiments. GFP-LC3 mice were from Riken with consent on their use from Dr. Mizushima, Noboru in Tokyo Medical and Dental University (10). Mice were fed with standard lab. chow and water ad libitum. The animals were maintained in the KNU animal facility at 20~22℃ under 40~60% relative humidity and a 12 h/12 h (light/dark) cycle for at least 7 days prior to the experiment. All experiments were approved by the Institutional Animal Care and Use Committee of Kangwon National University. In addition, the ethical guidelines described in the KFDA Guide for the Care and Use of Laboratory Animals was followed throughout the experiments.
Tumor cell lines and tumor models
Mouse colon carcinoma CT26 (American Type Culture Collection, Manassas, VA), and human Her-2/neu expressing CT26 (Her-2/CT26) were used (11). To establish subcutaneous (s.c.) tumors, 2×105 Her-2/CT26 tumor cells were injected on the left flank of BALB/c mice. The tumor growth was measured by caliper 3 times a week.
DC vaccine and OT-I proliferation assay
Bone marrow-derived DC was prepared as described previously (12). Bone marrow cells were incubated with 10 ng/ml of GM-CSF for 7 days and BM-DC were incubated with Ovalbumin (OVA, 100 ng/ml) for 18 hrs. These cells were further incubated with α-Mangostin (1 uM) for four hours before i.v. injection into mice adoptively transferred with carboxylfluorescein diacetate succinimidyl ester (CFSE)-labeled OT-I cells. To prepare CFSE-labeled OT-I cells, OVA-specific CD8+ T cells (>90% of which were Vα2-positive) were isolated from OT-I mice using magnetic beads. These cells were labeled with 5 uM CFSE and i.v. transferred into their syngenic C57BL/6 mice. Two days after DC vaccination, lymphoid cells from the lymph nodes or spleen of the recipient mice were stained with phycoerythrin (PE)-conjugated anti-V α2 and anti-CD8 antibodies and then analyzed by flow cytometry.
Antibodies
All Abs for flow cytometry were from BD Biosciences (San Jose, CA) including anti-CD8-APC and anti-Vα2-PE Ab. For the western blotting, anti-phospho-eIF2α, anti-β-actin and anti-rabbit/mouse HRP antibodies (Cell Signaling Technology) were used.
Confocal microscopy for autophagy activation
GFP-LC3 mice were sacrificed, and the small intestine was dissected and promptly washed with PBS. The tissue was fixed in formalin for 18 hours. Fixed tissue was embedded in OCT and sectioned on a cryotome into 8µm sections. Slides were washed with PBS and directly applied to attach coverglass. Images of the sections were collected using Zeiss confocal system (13).
Endoplasmic reticulum assay
To assess cellular ER stress, we checked the splicing of XBP1, the target of ER stress sensor kinase IRE1α, which was well-known to be activated by ER stress (14). We used the CT26 cells, and transfected them with XBP1-venus DNA (kindly provided by Masayuki Miura, RIKEN, Japan) by lipofectamine 2000 (Invitrogen) following the manufacturer's instructions (14). To induce the ER stress in the CT26 cells, DMSO as negative control or thapsigargin (200 nM; Sigma) were added after plating cells in 96-well plate (2×104 cells/200µl/well) and serially diluted α-Mangostin or DMSO as negative control were added, respectively. After 24 hrs incubation at 37℃, 5% CO2, cells were lysed in digitonin buffer (50 mM Tris-HCl (pH7.5), 1 mM EDTA, 10 mM EGTA and 10µM digitonin) at room temperature for 30 min. We used the GEMINI EM (Molecular Device) to measure fluorescence of the spliced XBP1 (emission at 520 nm; excitation at 485 nm).
Western blot
CT26 cells were cultured and treated with thapsigargin (200 nM) in the presence of DMSO or α-Mangostin. Western blot analysis was performed using mAbs to phosphor-eIF2α or β-actin as previously reported (14).
Preparation of α-Mangostin
The pericarps of G. mangostana L. were collected in Indonesia. The separation and identification of α-Mangostin was conducted as described previously (15). The purity of α-Mangostin was determined to be >95% based on HPLC-UV analysis.
RESULTS
Administration of α-Mangostin inhibited the growth of transplanted colon cancer in vivo
α-Mangostin is a xanthon derivative contained in the fruit hull of mangosteen (Garcinia mangostana L.), and was reported to induce apoptotic effect with Thapsigargin-like Ca2+-ATPase inhibitory effect (16). Likewise, α-Mangostin induced apoptosis in several human cancer cells (17-19). However, recently it was reported that α-Mangostin induced autophagic cell death, but not apoptosis in some brain cancer cell lines as assessed by DNA ladder fragmentation and caspase activities (1). Based on the previous reports supporting the anti-tumor effect of α-Mangostin, we decided to access whether oral administration of α-Mangostin can influence on the growth of colon cancer cells in mouse transplant model. To do this, groups of mice were subcutaneously injected with 2×105 cells of colon cancer cells, CT26 expressing Her-2/neu (Her2/CT26 cells), and orally administered with 20 mg/kg of α-Mangostin daily starting one day after tumor challenge. As expected, mice treated with α-Mangostin have significantly smaller tumor mass than control mice treated with DMSO (Fig. 1), supporting previous report showing anti-tumor effect of α-Mangostin as tested against Her2/CT26 cells.
α-Mangostin induced autophagy activation in vivo
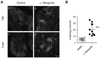
Although autophagy induction by α-Mangostin in several cell lines were shown previously (1), there was no report showing the influence of α-Mangostin on autophagy activation in vivo. Thus, to assess the influence of orally administered α-Mangostin on autophagy activation in vivo, GFP-LC3 transgenic mice, which have been widely used to detect autophagosome by LC3-II dot formation (10), was adopted. GFP-LC3 transgenic mice were orally administered with 20 mg/kg of α-Mangostin daily for three days and GFP puncta formation by LC3-II conversion in small intestinal villi and crypt epithelial cells including goblet cells and Paneth cells were monitored by confocal microscopy at 18 hrs after final administration (Fig. 2). α-Mangostin induced autophagic activation as determined by LC3-II conversion mainly in Paneth cells as well as in Goblet cells, suggesting that orally administered α-Mangostin induced autophagy activation in small intestinal epithelial cells in vivo.
α-Mangostin did not increase Ag-specific CTL priming by dendritic cells
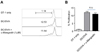
Autophagy-mediated activation of dendritic cell (DC) was known to be essential for a successful Ag presentation (20), especially for cross-presentation of Ag (21). Thus we accessed whether α-Mangostin would increase the cross-priming of CD8+ T cells by DC. To do this, bone-marrow derived DCs pulsed with OVA protein (0.1 mg/ml) were prepared, and incubated for four hrs with α-Mangostin (1µM) or DMSO as control prior to i.v. transfer into mice (5×105 cells/mouse) following adoptive transfer with CFSE-labeled OT-I CD8+ T cells. To monitor the effect of α-Mangostin, the dilution of CFSE on OT-I T cells by DC/OVA or DC/OVA/α-Mangostin was compared at 3 days after DC vaccination (Fig. 3). Despite the importance of autophagy on Ag presentation, activation of autophagy by α-Mangostin did not significantly increase OVA-specific T cell proliferation, suggesting that the antitumor activity by α-Mangostin might not be due to the augmentation of tumor Ag-specific CTL responses.
α-Mangostin reduced ER stress in vitro
Recently it was suggested that ER stress caused by accumulation of unfolded protein in the ER was associated with a wide range of diseases including cancer, cardiac disease, neurodegenerative diseases, and intestinal inflammation (22). Although α-Mangostin was also reported as SAECA inhibitor like thapsigargin, a well-known ER stress inducer (4,16), we wondered whether the treatment of α-Mangostin on thapsigargin-treated cells would influence on ER stress in vitro. In this regard, autophagy activation was recently reported to ameliorate ER stress (23), and autophagy activation by α-Mangostin was reported to possibly ameliorate ER stress induced by thapsigargin treatment.
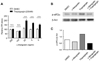
We used ER stress reporter system, which expresses fluorescence by ER stress-induced XBP1 splicing (14), to monitor cellular ER stress. Mouse colon cancer CT26 cells were transfected with XBP1-venus reporter, and incubated with 200 ng/ml of thapsigargin for 4 hrs in the presence or absence of the different concentration of α-Mangostin. The treatment of CT26 cells with 200 nM of thapsigargin induced XBP1 splicing monitoring by GFP expression, suggesting ER stress induction (Fig. 4A). Interestingly, ER stress-induced XBP1 splicing was significantly reduced by co-treatment of cells with 100 ng/ml and 250 ng/ml of α-Mangostin. These results suggest that the treatment of cells with α-Mangostin can reduce ER stress caused by thapsigargin.
Increased eIF2α phosphorylation by ER stress was ameliorated by α-Mangostin
To confirm the reduction of thapsigargin-mediated ER stress by α-Mangostin, CT26 cells were treated with 200 ng/ml of thapsigargin with or without 250 ng/ml of α-Mangostin and assessed phosphorylation of eIF2α by western blot analysis. PERK-eIF2α is another UPR pathway activated by ER stress (Fig. 4B and C). Treatment of cells with thapsigargin increased eIF2α phosphorylation, but α-Mangostin alone did not. Consistent with reduced XBP1 splicing in thapsigargin treated CT26 cells after α-Mangostin treatment, α-Mangostin reduced eIF2α phosphorylation by thapsigargin, suggesting that α-Mangostin inhibits ER stress induction by thapsigargin.
ER stress with autophagy blockade accelerated tumor growth in mouse colon cancer model
Previously, it was reported that thapsigargin specifically inhibited autophagy flow by blocking autophagosome fusion with lysosome (3). It is quite interesting because both thapsigargin and α-Mangostin were known as inhibitors for SERCA, and they block the pumping of calcium into the sarcoplasmic and endoplasmic reticulum and result in ER stress induction (4,16). As shown in Fig. 4, α-Mangostin did not induce ER stress in the concentration we used, whereas thapsigargin significantly increased ER stress. To assess the consequence of coadministration of α-Mangostin and thapsigargin on tumor growth, groups of mice were subcutaneously injected with 2×105 cells of Her2/CT26 cells, and α-Mangostin and thapsigargin was administered every day starting one day before tumor injection (Fig. 5). When the growth of tumors was monitored, we found that the growth retardation observed in mice treated with α-Mangostin alone disappeared in mice coadministered with thapsigargin. On the contrary, coadministration of α-Mangostin with thapsigargin rather increased tumor growth as compared with DMSO-treated mice. These result suggested that autophagic arrest by systemic treatment of thapsigargin in vivo inhibited antitumor activity of α-Mangostin.
DISCUSSION
Autophagy is a cellular process for the removal of abnormal intracellular organelles, pathogens and the recycling of cytoplasmic material to obtain energy (24). This pathway also has been involved in immune responses, including antigen processing and presentation pathway (20,21,24,25). Autophagy mediated intracellular lysosome-dependent degradation of abnormal cellular macromolecules and pathogens, and resulted in the accumulation of autophagolysosomes in the cytosol (24). During the formation of autophagolysosome, LC3 and ATG12, two ubiquitin-like protein systems, play an important role for the autophagosomal membrane formation and expansion (26). The conversion of soluble LC3-I to the membrane bound LC3 II is often used to monitor autophagy activation in the cell and GFP-LC3 constructs have been used successfully to track autophagy activation in vivo and in vitro (26). In the current study, we adopted GFP-LC3 mice to monitor whether oral administration of α-Mangostin would induce autophagy activation, and the increased GFP dots in intestinal epithelium suggesting the activation of autophagy.
The unfolded protein response (UPR) is an evolutionary conserved process to ER stress induced by accumulation of misfolded proteins in ER lumen (27). UPR pathway is known to be important to the development and survival of cells, especially for highly secretory cells or cells that synthesize large quantities of proteins (27). In mammals, the UPR is mediated by three transmembrane proteins that reside in the ER (27): PKR-like ER-resident kinase (PERK), activated transcription factor 6 (ATF6) and inositol requiring enzyme 1 (IRE1) pathways. The accumulation of misfolded proteins within the ER attracts the chaperone protein, Grp78 and activates UPRs to facilitate protein folding, attenuate translation, and activate ER-associate degradation (ERAD) pathway. Briefly, ER stress-mediated dissociation of PERK from Grp78 leads to transphosphorylation of PERK and subsequently increases phosphorylation of eIF2α, which attenuate translation of CAP-dependent protein. At the same time, phosphorylated eIF2α induces transcription of ATF4 which activates a number of UPR genes including Grp78, CHOP and GADD34. ATF6 upon release from a Grp78 translocates to the Golgi and then induces transcription of several UPR genes including X-box protein 1 (XBP1), CHOP and GRP78. The most evolutionary conserved of the UPRs is IRE1. Activated IRE1 can splice the Xbp1 mRNA into the active form of Xbp1 (XBP1s) which regulates important genes involved in ER translocation, expansion and ERAD activation. Activated IRE1 also has kinase activity, and signals through TRAF2 that in turn activates JNK and its downstream targets that could promote inflammatory response and/or apoptosis. In the current study, we used XBP-1 reporter which express GFP fluorescence upon XBP-1 splicing by IRE-1 during ER stress. In addition, we detected phosphorylation of eIF2α to assess alteration of ER stress status in cells treated with thapsigargin and α-Mangostin. Consequently, we found that ER stress induced by thapsigargin can be reduced by cotreatment of α-Mangostin. In addition, although α-Mangostin was shown to inhibit SAECA like thapsigargin, it did not induce ER stress under concentration we used. Since thapsigargin specifically inhibits the fusion of autophagosome with lysosome, the last step in autophagic pathway (3), it might induce the accumulation of ER stress by the defect in autophagic elimination of ER stress.
Interestingly, recent studies suggested that UPR mediated by ER stress was known to be linked with autophagy activation (6,7). The activation of autophagy pathway by ER stress includes the following pathways: IRE1-mediated JNK activation and PERK/eIF2α phosphorylation (6,7). Although both of UPR and autophagy are important for cellular homeostasis, unfolded protein responses triggered by ER stress have been shown in several tumors, particularly in area with hypoxia, and they help tumor cells survive in stressful tumor microenvironment (8). Thus, the outcome of autophagy activation by small molecule drugs inducing autophagy activation on the survival of tumor was difficult to predict. Indeed, there was controversy about the pro-apoptotic or anti-apoptotic roles of autophagy (6,21). Further, in several recent experiments done by others, it was suggested that ER stress can induce autophagy and the induction of autophagy by ER stress is beneficial to relieve ER stress and consequently ameliorate the disease progress (23). However, the relationship between the two processes has not been fully elucidated yet. In the current study, activation of autophagy by α-Mangostin significantly inhibited tumor growth, suggesting it exerted proapoptotic function against tumor cells. Likewise, thapsigargin, a well-known ER stress inducer, which was recently proven to block autophagy flow, completely blocked the anti-tumor activity by α-Mangostin. Collectively, these results suggest that autophagy activation by α-Mangostin might be important for the successful antitumor activity.


 XML Download
XML Download