

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
NK
natural killer
TNF-α
tumor necrosis factor-α
IFN-γ
interferon-γ
ADCC
antibody-dependent cellular cytotoxicity
LAD
leukocyte adhesion disorder
HSV
herpes simplex virus
HED-ID
hypohidrotic ectodermal dysplasia with immune deficiency
XLP
X-linked lymphoproliferative disease
SLAM
signaling lymphocyte activation molecule
ITIM
immunoreceptor tyrosine-based inhibition motif
ITAM
immunoreceptor tyrosine-based activation motif
SHP-1
Src homology 2 domain-containing phosphatase 1
NCRs
natural cytotoxicity receptors
LAT
linker for activation of T cells
NTAL
non-T-cell activation linker
YINM
tyrosine-based signaling motif
ITSM
immunoreceptor tyrosine-based switch motif
GEF
guanine nucleotide exchange factor
SLP-76
SH2 domain-containing leukocyte phosphoprotein of 76 kD
INTRODUCTION
Natural killer (NK) cells are derived from bone marrow-derived lymphocytes and belong to the innate immune system. In contrast to T and B cells, activation of NK cells is controlled by germ line-encoded receptors that do not undergo somatic DNA recombination and mutation. NK cells can provide early immune defense against virus-infected and transformed cells. Upon activation, NK cells eliminate target cells by polarized exocytosis of cytotoxic granules containing perforin, granzymes as well as Fas ligand (1). They also control immune responses by secretion of cytokines and chemokines including tumor necrosis factor-α (TNF-α) and interferon-γ (IFN-γ). NK cells via these effector functions play important roles in innate resistance to viral pathogens, tumor surveillance, shaping adaptive immunity, graft-versus-leukemia activity, antibody-independent natural cytotoxicity and antibodydependent cellular cytotoxicity (ADCC) against IgG-coated target cells.
The important role of NK cells in human health has been underscored by a wide spectrum of disease associations with NK cell dysfunction. Patients with leukocyte adhesion disorder (LAD), a syndrome due to the deficiency of β2 integrin subunit, display attenuated ADCC and natural cytotoxicity by NK cells (2,3) and suffer severe recurrent infections of bacteria, infections of herpes simplex virus (HSV), and impaired immunity (4-6). NK cells from patients with hypohidrotic ectodermal dysplasia with immune deficiency (HED-ID) have defective natural cytotoxicity but not ADCC (6). In NK cells from patients with X-linked lymphoproliferative disease (XLP), activating receptors such as the signaling lymphocyte activation molecule (SLAM) family member 2B4 (CD244) and NTB-A induce inhibitory rather than activating signals as essential signaling adaptor SAP is deficient. Thus, 2B4 or NTB-A-mediated autologous killing of Epstein-Barr virus-infected B cells is abolished in XLP patients (7,8).
Recently, NK cells have gained increasing attention and are now being considered as promising therapeutic tools for cancer immunotherapy due to their intrinsic ability to rapidly recognize and kill cancer cells, while sparing normal healthy cells. Allogeneic NK cells exhibit potent anti-tumor activities that are beneficial in the setting of hematopoietic stem cell transplantation into patients with acute myeloid leukemia (9). Moreover, KIR-HLA-incompatible NK cells were shown to lyse melanoma and renal cell carcinoma cells in vitro (10). Moreover, administration of rituximab (anti-CD20) to patients with large-cell non-Hodgkin's lymphoma induced NK cell cytotoxicity in vivo via ADCC (11).
Despite significant progress made on the role of NK cells as a pivotal sentinel in virus and tumor surveillance, the molecular mechanisms that regulate NK cell responses remain unclear, which restricts the use of NK cells as a therapeutic measure. NK cells possess a combinatorial array of activating and inhibitory receptors instead of a dominant single antigen receptor to sense their target cells. Whether NK cells are activated toward target cells or maintain tolerance to self is determined by the signal balance between activating and inhibitory receptors (12-14). Thus, understanding how signals from discrete activating receptors cooperate to control NK cell activation and how signals from inhibitory receptor antagonize such activation may provide strategies for harnessing NK cells in various clinical settings.
ACTIVATING RECEPTORS AND INTRACELLULAR SIGNALING
A large array of NK cell activating receptors that are structurally distinct has been characterized (15,16). In contrast to inhibitory receptors that display variegated expression on NK cells, activating receptors are uniformly expressed on most NK cells. Furthermore, activating receptors trigger diverse and discrete signaling cascades, whereas inhibitory receptors induce a common signaling mechanism where their cytoplasmic immunoreceptor tyrosine-based inhibition motif (ITIM) recruits the phosphatase Src homology 2 domain-containing phosphatase 1 (SHP-1) to dephosphorylate signaling molecules in the activation of NK cells (17,18).
Among activating receptors containing immunoreceptor tyrosine-based activation motif (ITAM) or associated with ITAM-carrying adapters are the low-affinity Fc receptor FcγRIIIa (CD16), natural cytotoxicity receptors (NCRs) such as NKp30 (CD337), NKp44 (CD336), and NKp46 (CD335), activating KIRs including KIR2DS and KIR3DS, and CD94/NKG2C (CD94/159c) heterodimer (16). Upon engagement, these ITAM-associated receptors are tyrosine-phosphorylated by Src family kinases, and the phosphorylated ITAM-bearing subunits recruit Syk and ZAP-70 tyrosine kinases. Thereafter, these kinases phosphorylate linker for activation of T cells (LAT) and non-T-cell activation linker (NTAL) leading to activation of PI3K, PLC-γ1, 2 and Vav 2, 3 (19-23).
Non-ITAM receptors include C-type lectin-like family member NKG2D (CD314), CD2, 2B4, CRACC (CD319), NTB-A, and the immunoglobulin-like type I transmembrane glycoprotein DNAM-1 (CD226). Upon stimulation, NKG2D associates with the adaptor protein DNAX-activating protein of 10 kD (DAP10) through its transmembrane region (24). The cytoplasmic tail of DAP10 carries a tyrosine-based signaling motif (YINM). This YINM, after phosphorylation by Src family kinases, recruits either a p85 subunit of PI3K or the small adaptor protein Grb2. Both interactions can activate Vav1 and PLC-γ2 and thus are essential for the activation of Ca2+ mobilization and cytotoxicity toward target cells (25). Unlike ITAM-associated receptors, NKG2D signaling is independent of Syk, ZAP70, LAT and NTAL (26-28). SLAM family receptors including 2B4, NTB-A, and CRACC are also phosphorylated by Src family kinases at immunoreceptor tyrosine-based switch motifs (ITSM) in their cytoplasmic domain (29). This ITSM, after phosphorylation, recruits small adaptor proteins such as SAP, EAT-2, and ERT (30-33). SAP then binds Src family kinase Fyn and this SAP-Fyn interaction is indispensable for 2B4-mediated NK cell activation (34,35). Vav1, PLC-γ2, and c-Cbl are phosphorylated by 2B4 stimulation and implicated in such activation (31,36). DNAM-1 signaling in NK cells remains largely unclear. Cytoplasmic tail of DNAM-1 can be phosphorylated by Src-family kinases and protein kinase C, and recruit actin-binding proteins (37,38).
Although various signaling molecules implicated in NK cell activation have been uncovered, it has been difficult to identify critical molecules essential for NK cell activation because NK cells use diverse kinases, adaptors, and signaling modules depending on the engagement of different activating receptors. However, intersection of signals from discrete activating receptors by a single class of ITIM-associated inhibitory receptors implies that there exist critical control elements in the signaling pathway of NK cell activation. Such examples include PLC-γ and Vav proteins that play a non-redundant role in NK cell activation by multiple activating receptors, and their deficiency is manifested by functional defects of NK cells (20,39,40).
Src family kinases that include Lck, Fyn, Src, Lyn, Yes, and Fgr are important for NK cell activation as most of the activating receptors on NK cells are dependent on their activities for activation. However, no single Src family kinase is indispensable because their function is highly redundant for NK cell activation (41,42). PLC-γ activation that induces Ca2+ mobilization from endoplasmic reticulum is critical for exocytosis of lytic granules and cytokine production. Although PLC-γ1 and PLC-γ2 are redundant in ITAM-dependent signaling, NKG2D/DAP10 and 2B4 signaling are highly dependent on PLC-γ2 (36,40).
Vav has three isoforms, Vav1, Vav2, and Vav3, and NK cells express all of these isoforms (20). Vav acts as a guanine nucleotide exchange factor (GEF) that activates small GTPase proteins of the Rho family and as an adaptor protein through C-terminal SH2 and SH3 domains (43-45). It has an important role in the actin cytoskeleton dynamics for the co-activation receptor clustering and the lytic granule polarization, as well as in the lymphocyte receptor signaling including PLC-γ activation (45,46). Vav1 is essential for DAP10 signaling, and Vav2 and Vav3 are required for ITAM-associated receptors (20,47,48). Thus, the identification of critical signaling molecules that function as essential checkpoints for NK cell activation provides clues for understanding the mechanism of cooperative interplay by multiple co-activation receptors.
SYNERGISTIC ACTIVATION OF NK CELLS
Activation of NK cells is tightly controlled by the requirement for the engagement of multiple co-activation receptors on NK cells upon encounter with target cells (16,49). The combined signals from specific pairs of NK cell receptors, such as NKG2D and 2B4, can operate in synergy to induce potent cytotoxicity toward sensitive target cells (16,50), while other combinations can result in different outcomes: no enhancement over each activating signal or simple additive effect. The mechanisms how distinct signals from different co-activation receptors are integrated to achieve proper functional activity are still unclear and difficult to study because each co-activation receptors use distinct signaling modules. Among the combinations of activating receptors, NKG2D and 2B4 as well as 2B4 and DNAM-1 are able to provide synergistic activation in resting NK cells, whereas NKG2D and DNAM-1 do not synergize. Even though NK cells are activated by synergistic signals from multiple co-activation receptors, inhibitory signals from a single class of ITIM-containing receptors, such as KIRs and CD94-NKG2A, are dominant over synergistic activating signals. This suggests that there are central common checkpoints where signals from multiple co-activation receptors are converged and integrated but inhibitory signals intersect.
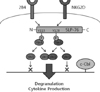
A study with NKG2D and 2B4 synergy advocates such an example of checkpoint and an elaborate mechanism for regulating synergistic activation (36). The central role of Vav1 in the signaling pathway for NK cell activation is emphasized by its identification as an essential target of SHP-1 recruited to MHC class I-specific inhibitory receptors (51,52). The Vav1 is phosphorylated by the engagement of NKG2D, 2B4 and DNAM-1 on NK cells and functions as an essential component in signaling from such activating receptors. Synergistic activation of NK cells via NKG2D and 2B4 combination results in synergistic phosphorylation of PLC-γ2, Ca2+ mobilization and cytotoxic degranulation, which are completely dependent on Vav1. Signals from single receptor activation can also induce Vav1 phosphorylation, but not the subsequent events such as the phosphorylation of PLC-γ2, Ca2+ mobilization, degranulation and cytokine secretion. Such dichotomy of NK cell activation can be explained by the finding that single receptor signal is unable to overcome activation threshold shaped by the E3 ubiquitin ligase c-Cbl mediating the inhibition of Vav1-dependent activation. Such threshold can be overcome by the convergence of signals from NKG2D and 2B4 to achieve optimal activation of Vav1 (Fig. 1). This mechanism supports the presence of a central checkpoint for NK cell activation where Vav1-mediated activation and c-Cbl-mediated inhibition compete with each other.
Consecutive study of synergy between NKG2D and 2B4 revealed another interesting mechanism for synergy (53). The phosphorylation of Vav1 by NKG2D and 2B4 synergy is equivalent to the sum of the extent of phosphorylation induced by each receptor alone. In contrast, phosphorylation of Vav1 was not augmented by inactive receptor combination although each receptor induced Vav1 phosphorylation independently. Thus, synergy involves the integration of different activation signals for optimal activation of Vav1, but such requires combination of specific receptor pairs, suggesting the presence of an upstream regulatory element to control Vav1 activation. The adaptor proteins SH2 domain-containing leukocyte phosphoprotein of 76 kD (SLP-76) and LAT are known to contribute to Vav1 signaling and are essential components of forming signaling complexes in T cells (54,55). Furthermore, the activation of PLC-γ, which is required for synergistic activation of NK cells (36), is dependent on both SLP-76 and LAT in T cells (56-58). In NK cells, the synergistic signaling pathway induced by co-engagement of NKG2D and 2B4 is largely dependent on SLP-76, but not LAT. Signal from NKG2D induces preferential phosphorylation at tyrosine (Tyr)128 of SLP-76, whereas signal from 2B4 induces phosphorylation of SLP-76 at Tyr113 (Fig. 1). Both tyrosine phosphorylations at Tyr113 and Tyr128 of SLP-76 are required for the synergistic activation of NK cells (53). The SLP-76 phosphorylation in NK cells is differentially regulated compared to TCR signaling in T cells because the single TCR engagement is sufficient for phosphorylation of both tyrosine residues in SLP-76 (54). Moreover, complementary phosphorylation of SLP-76 at N-terminal tyrosine residues is a property of synergizing receptors that induce natural cytotoxicity because the engagement of the Fcγ receptor CD16 also induces SLP-76 phosphorylation at both tyrosines. Selective phosphorylation of SLP-76 at Tyr113 or Tyr128 each enabled the binding of SLP-76 to Vav1 to promote Vav1 phosphorylation. Thus, SLP-76 may serve to integrate signals from synergizing co-activation receptors by recruiting Vav1 to its phosphorylated tyrosine residues. This suggests that permissive activation of NK cells is kept in check by the requirement for the complementary phosphorylation of SLP-76 during synergy among receptors that stimulate natural cytotoxicity.
CONCLUDING REMARKS
NK cell activation upon encounter with target cells is a result from dynamic process of signaling cascades triggered by combination of multiple receptors on NK cells. Thus, diverse mechanisms may control each step of NK cell activation depending on the signaling pathways originated from not only a single receptor but also a cooperation of co-activation receptors. In this review, the requirement for complementary phosphorylation of SLP-76 by synergizing receptors for optimal activation of Vav1 and thereby to overcome inhibition by c-Cbl is introduced as recently discovered mechanism regulating synergistic activation of NK cells. A challenge for future study is to find different checkpoints and signaling mechanisms that control synergistic activation of NK cells. This information will provide innovative and applicable strategies for exploiting NK cells in clinical immunotherapy.
 XML Download
XML Download