

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
T cell immunoglobulin mucin domain (TIM)-3 is expressed in various leukocyte subpopulations and modulates their functions (1). It is expressed on the surfaces of exhausted T cells and involved in the downregulation of effector function of T cells (2-4). TIM-3 on dendritic cells promotes the uptake of apoptotic cells via interaction with phosphatidylserine but suppresses the immunogenicity of nucleic acid via interaction with HMGB1 (5,6). TIM-3 expression in mast cells influences cytokine production and apoptosis of these cells (7). However, the regulation of TIM-3 expression has not been well known.
Mast cells play a significant role in various immune responses. These cells lead allergic symptoms through secretion of mediators and cytokines upon cross-linking of FcεRI (8). Mast cells contribute defense against pathogen; mice devoid of mast cells die from infection that does not result in death in wild type mice (9). Protective immunity is enhanced by mast cell production of TNF-α and recruitment of neutrophils to the infectious site (9). Mast cells are also required for the development of collagen-induced rheumatoid arthritis and the induction of tolerance to skin graft (10,11).
To understand the regulation of TIM-3 expression in mast cells, we investigated the effect of various cytokines on TIM-3 transcription and the activity of TIM-3 promoter in relation with TGF-β1 stimulation of mast cells.
MATERIALS AND METHODS
Reagents
IFN-α, IFN-λ, TGF-β1, IL-10, and TNF-α were purchased from R&D Systems (Minneapolis, MN, USA). Expression vectors for Smad2 and Smad4 were kindly provided by Dr. Cho (Dept. Biochemistry, Ajou University School of Medicine, Korea).
Cell culture
HMC-1, a human mast cell line, was kindly provided by Professor Hyung Min, Kim (Kyunghee University, Suwon, Korea). HMC-1 cells were maintained in IMDM supplemented with 10% FBS, penicillin-streptomycin (each 100 U/ml, 100 ug/ml). Similarly, HEK293 cells were maintained in RPMI 1640 (Gibco BRL, Paisley, Scotland) supplemented with 10% FBS.
Real-time RT-PCR
The total RNA was isolated using RNA STAT-60 (Tel-Test, IN C., Friendwood, TX, USA) and reverse transcribed using Superscriptase II (Invitrogen, Carsbade, CA, USA). Real-time PCR was performed using primers (5'-TCCAAGGATGCTTACCACC AG-3': 5'-GCCAATGTGGATATTTGTGTTAGATT-3') and a Taq Man probe (5'-ACATGGCCCAGCAGAGACACAGACACT-3') for TIM-3 transcript which was normalized to GAPDH transcript levels.
Plasmid construction
Luciferase reporter vectors were constructed by ligation of human TIM-3 promoter region DNA fragment into pGL-Basic vector (Promega, Madison, WI, USA). Human TIM-3 promoter DNA was amplified by PCR using genomic DNA isolated from HMC-1 cells. For amplification of TIM-3 -1677~+144 DNA fragment, primers TIM3+1 (5'-GGAGCTTGCAGAAGAAAAGTCAGAGGACACCTCTGTTAGG-3') and 5'-AGAGCCTTGACCAAGTTCATGCTGCTAATAAAAATAACCCCAG-3' were used. For TIM-3 -872~+144 DNA fragment, primers TIM3+1 and 5'-CTTTTGCTTTTAAGGTGTCCAGATAAAGGTCACACTCCCAG-3' were used. For TIM-3 -349~+144 DNA fragment, primers TIM3+1 and 5'-CTGTGACCAAAGTTTATGAAGCC-3' were used. The PCR products was cloned into Topo TA cloning vector (Invitorgen) and the nucleotide sequences were verified by comparison with the gene sequence (NW_001838954). Then TIM-3 promoter DNA was subcloned into pGL-Basic vector using Nhe I and Bgl II and designated as T3U(1.8)-luc, T3U(1.0)-luc and T3U(0.5)-luc.
Luciferase reporter assay
HMC-1 cells (1×106) were transfected with 3.6µg of luciferase reporter vector together with 400 ng of pEGFP-N1 plasmid (Clonetech, Mountain View, CA, USA) using electroporator (Digital Bio Technology, Seoul, Korea). The cells were incubated for 48 h in 5% CO2 incubator at the 37℃, and then luciferase activity was analyzed using luciferin (Promega, Madison, WI, USA) and luminometer (Molecular devices, Sunnyvale, CA, USA). The transfection efficiency was analyzed by GFP-expressing cell frequencies using flow cytometer (FacsCanto). Similarly, luciferase activity was assessed in HEK293 cells transfected with DNA using Lipofectamin 2000 (Invitrogen, Carsbade, CA, USA).
RESULTS
TIM-3 transcription was upregulated in HMC-1 cells by TGF-β1 stimulation
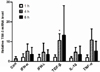
In recent studies, TIM-3 mRNA and protein expression levels were up regulated in TGF-β1 stimulated-human mast cells (12) but the transcriptional regulation of TIM-3 by other stimulants has not been well explored. To address this issue a human mast cell line, HMC-1 cells were treated with various cytokines for the indicated time and then TIM-3 mRNA levels were determined by real-time RT-PCR. TIM-3 mRNA expression was not significantly enhanced by treatment with the indicated concentration of IFN-α IFN-λ, IL-10, or TNF-α (p>0.05) but was significantly increased by TGF-β1 stimulation of HMC-1 cells for 4 h (p<0.005) (Fig. 1).
TIM-3 promoter activity was increased in HMC-1 cells by TGF-β1 stimulation
To know the TIM-3 promoter is responsive to TGF-β1 stimulation, we first examined the basal activity of TIM-3 promoter by luciferase reporter assay using vectors depicted in Fig. 2A. Luciferase activities in both HEK293 cells and HMC-1 cells transfected with T3U(0.5)-luc that contains proximal TIM-3 promoter were significantly higher than that of controls transfected with the empty vector pGL-Basic (Fig. 2B and C). Also, luciferase activities driven by T3U(1.0)-luc and T3U(1.8)-luc, respectively were significantly higher than that of controls but lower than that driven by T3U(0.5)-luc. These results indicate that the proximal TIM-3 promoter spanning from +144 to -349 may consist of sufficient element for the basal level transcription of TIM-3 in HMC-1 cells.
We next analyzed the TIM-3 promoter activity in the presence of TGF-β1 stimulation (Fig. 3). Luciferase activity driven by each luciferase reporter vector in the presence of TGF-β1 was presented as fold induction relative to that in the absence of TGF-β1. Luciferase activity in HMC-1 cells transfected with T3U(0.5)-luc or T3U(1.8)-luc was significantly increased (2.3 and 2.6 fold, respectively) by TGF-β1 stimulation compared to that in the absence of TGF-β1 stimulation (p<0.001). However, luciferase activity in HMC-1 cells transfected with T3U(1.0)-luc or the empty pGL-Basic was not significantly increased by TGF-β1 stimulation compared to that in the absence of TGF-β1 stimulation (p>0.01). These results suggest that TIM-3 promoter region may contain the TGF-β1 responsive elements.
TIM-3 promoter activity was upregulated by Smad2 and Smad4 overexpression
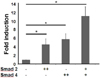
To support the TGF-β1 responsiveness of the TIM-3 promoter, we assessed TIM-3 promoter activity in the cells overexpressing Smad2 and Smad4 that were reported to be involved in the gene expression induced by TGF-β1 (13,14). Given that TGF-β responsive promoter activity was increased by overexpression of either Smad2 or Smad3 in HEK293 cells even in the absence of TGF-β stimulation (15), we analyzed TIM-3 promoter activity in HEK293 cells overexpressing Smad2 and/or Smad4 without TGF-β1 treatment (Fig. 4). Compared to control, luciferase activity was significantly increased in HEK293 cells by overexpression of Smad2, Smad4 or both (4.5, 5.8 and 11 fold, respectively) (p<0.01). These results imply that TIM-3 promoter region may respond to TGF-β1 stimulation through Smad2 and Smad4 involvement.
DISCUSSION
In this study, we revealed that TIM-3 mRNA expression in a human mast cell line was increased by TGF-β1 stimulation but not by other stimuli such as interferon α and λ, TNF-α, and IL-10. TGF-β1 affects mast cell survival and functions. TGF-β1 inhibits IL-3-dependent mast cell proliferation and counterbalances the effect of IL-4 on mast cell survival, migration, and FcεRI expression (16,17). Also TGF-β1 can elicit mouse mast cell protease-1 expression (18) and mast cell tryptase expression in experimental emphysema model (19). Furthermore, TGF-β1 in both soluble and regulatory T cell-surface bound forms can escalate IL-6 production by mast cells (20). Our results (21) and the previous report by Wiener et al.(12) add another one to the effects of TGF-β1 on mast cells i.e. induction of TIM-3 expression. Except the report by Nakae et al.(7) that TIM-3 cross-linking by an anti-TIM-3 polyclonal Ab can promote IL-4, IL-6 and IL-13 production but suppress mast cell apoptosis, little information is currently available regarding the role of TIM-3 in mast cell function.
We also demonstrated that TIM-3 promoter -349~+144 bp region relative to the transcription start site was crucial for the basal and TGF-β1-induced TIM-3 transcription in HMC-1 cells. Since T3U(0.5)-luc driven luciferase activity in HMC-1 cells was greater than that driven by T3U(1.8)-luc or T3U(1.0)-luc, the DNA fragment of -349~+144 bp seems to contain elements for basal transcription of TIM-3. Compatible to our results, Zhang et al. (22) reported that the basal TIM-3 promoter activity is localized to the region spanning -241 to + 63 bp in YT cells, a T/NK cell line. Apparently incompatible with our previous observation that TIM-3 promoter activity driven by the -1,362 to +144 bp region was not enhanced by TGF-β1 stimulation (21), luciferase activities under TIM-3 promoter -349~+144 bp region and -1,677~+144 was significantly increased in HMC-1 cells by TGF-β1 stimulation (Fig. 3). Interestingly, the luciferase activity under TIM-3 promoter -872~+144 was not significantly elevated by TGF-β 1 stimulation in the present study. Further study is required to investigate whether TIM-3 promoter -1,362 to -350 bp region may contain the element hindering the TGF-β1 responsiveness of TIM-3 promoter -349~+144 bp region.
We showed that overexpression of Smad2 and Smad4 upregulated TIM-3 promoter activity in HEK293 cells. Smad2 and Smad4 are downstream signaling molecules and transcription factors of TGF-β1 signaling (13,14). Although there are three predicted Smad binding elements in TIM-3 promoter -349~+144 bp region, further study should be followed to clarify whether Smad2 and Smad4 are directly bound to TIM-3 promoter or indirectly involved in TIM-3 transcription in TGF-β1 stimulated mast cells. In T cells, it was reported that two transcription factors regulated TIM-3 transcription in different modes. T-box transcription factor T-bet increases Tim-3 transcription via its interaction with Tim-3 promoter of mouse TH1 cells at approximately 400 bp upstream of the first ATG, whereas signal transducer and activator of transcription (STAT)-4 dose not bind to the Tim-3 promoter but reduces Tim-3 expression in murine T cells when it is knocked-out (23). At present, critical role of MEK, another downstream signaling molecule of TGF-β1 signal pathway, in TIM-3 induction is revealed in HMC-1 cells stimulated with TGF-β1 (21).
Conclusively, our results localize TIM-3 promoter activity in resting and TGF-β1 stimulated HMC-1 cells to the region from -349 to +144 bp relative to the transcription start site and propose a possible regulatory role of Smad2 and Smad4 in TIM-3 transcription of mast cells.


 XML Download
XML Download